


Keywords
Abstract
Membrane electron transfer underlies the central metabolic pathways for energy conversion. The photosynthetic and respiratory electron transport chains are complex apparatuses capable of generating a transmembrane proton gradient from sunlight or chemical energy. Recent exploitation of these apparatuses as energy convertors is of interest due to the availability and eco-friendliness of the biomaterial. Devices that utilize chemotrophic microorganisms to generate electricity have been known for over one hundred years. In these systems, called microbial fuel cells (MFC), one or more microorganisms catalyze charge transfer from a consumable substrate (acetate, glucose, etc.) to the electrode. Recently, MFCs based on phototrophic organisms have been actively developed. These devices, called photosynthetic microbial fuel cells (PMFC), still resemble the conventional MFC in that they also use living microbial cells to convert chemicals to electrical energy. However, the distinction between these two classes of fuel cells is that the MFC utilizes only the chemical energy of the organic substrate. At the same time, PMFCs are also capable of using solar energy. Common to both devices is the ability to utilize intrinsic electron transfer chains of bacterial metabolism as the primary mechanism of energy conversion. The widespread and accessible solar energy may permit PMFCs based on photosynthesis to become an inexpensive and efficient method for sunlight conversion. MFCs based on heterotrophs may be more promising in wastewater remediation and other ecological applications. This article reviews the latest advances in this field and emphasizes the remaining challenges.
The bibliography includes 205 references.
1. Introduction
Many primary energy-coupled metabolic pathways in living cells are based on the membrane-associated redox active complexes combined into electron transport chains (ETCs). These chains implement electron flow coupled with the transmembrane movement of protons. This process results in the generation of a transmembrane proton gradient. The proton gradient, in turn, activates ATP-synthase, a membrane enzyme responsible for ATP (adenosine triphosphate) synthesis from ADP (adenosine diphosphate) and inorganic phosphate.1,2 Microorganisms could synthesize ATP through generated transmembrane gradient more than 3 billion years ago.3,4 It is known that multiple forms of energy (electrical, osmotic, chemical, and, in the case of photosynthesis, solar) can be captured and transformed in the membranes of different biological ETCs. The elaborated mechanisms that nature uses to solve the energy problems of living organisms may ultimately also be used by humanity to solve the energy and ecological problems created by society. Interest in using these natural green processes is rapidly growing with increasing public concerns about emerging geopolitical and ecological risks for global energy needs.5,6
Systems that generate electricity or synthesize molecular hydrogen through the metabolism of microorganisms are called microbial fuel cells (MFC).7 Potter8 demonstrated approximately a century ago that heterotrophic bacteria and fungi could produce electricity in a two-electrode system. Considerably later, the concept of photosynthetic microbial fuel cells (PMFCs) has been evolving.9 In these systems, photosynthetic ETC of living phototrophic bacteria serves as the apparatus for electron generation instead of the respiration chain utilized in MFC. The primary energy source is sunlight for the PMFC instead of chemical bonds. The very high quantum efficiency of light-induced charge separation in photosynthetic reaction centers (RC) has attracted the attention of researchers.10,11 Biohybrid solar cells based on the isolated components of the photosynthetic apparatus are the alternative to the PMFC. These biohybrid cells differ from the PMFCs because they use subcellular pigment-protein-lipid complexes instead of integrating the entire phototrophic organisms alone or in a mixed community. The advantage of such a biohybrid system compared with PMFC is that the device has many fewer electron transfer steps and thus loses less energy to competing reactions and resistance of the electron transfer from ETC to the electrode. The disadvantage of such a biohybrid device is the absence of the native mechanism of repair and regeneration of the pigment-protein complexes and other electron transfer components found within the native membrane structures.12
This article focuses on various works exploring fuel cells based on organisms that implement heterotrophic and/or phototrophic metabolism. The second Section describes different scopes of such bio-hybrid devices. The third Section concerns the different electron transfer chains in nature with an emphasis on the respiratory and photosynthetic ETCs of bacteria and the photosynthetic chains of plants. These ETCs or components derived from them are the main driving force for all MFCs and the main components in many biohybrid devices. Understanding their structure and function is required to design effective microbial fuel cells or biohybrid solar cells. The following sections summarize the recently reported advances in the field of microbial fuel cells.
2. Practical application of biohybrid devices
ETC exploitation in biohybrid devices have a wide range of economic and environmental application (Fig. 1). The production of electricity or molecular fuel from the vital activity of bacteria or subcellular membrane complexes is a promising area of research because it is eco-friendly.13--15 Some MFCs can feed on organic waste in wastewater and thus be used for wastewater treatment.16 The ability to generate electricity from wastewater received worldwide attention when similar technology was introduced to various space programs worldwide when it was shown that MFCs could be used to solve the problem of human waste accumulation during extended space flights.7,17 Also, special MFCs can be used for seawater desalination and denitrification.18,19
![[{"id":"Y7xABIE6gX","type":"paragraph","data":{"text":" Practical application of electron transport chains"}}]](/storage/images/resized/Z9m0fHyyrjJjOKVwb44zBmx6Vd3GHUNQt8Hl4mmy_xl.webp)
PMFCs can work as biosensors. The photocurrent generated in PMFC depends on a number of factors, including the presence in the bacterial culture of chemical compounds that block membrane electron transfer. Certain pollutants, such as herbicides, act as inhibitors of photosynthetic electron transfer and attenuate the photocurrent dose-dependently. Due to this, the design of many biosensors of pollutants is based on the biohybrid electrode contained RCs, photosynthetic membranes, or intact photosynthetic microorganisms.20--23
The electron transfer stage of the metabolic pathways can be used for analytical purposes: special devices that measure the electrical signal from MFC can be applied to characterize the metabolic pathway. This method allows researchers to study single membrane redox enzymes and more complex and large structures from membrane fragments to microbial cells.24--27
3. The diversity of electron transfer chains
3.1. Photosynthetic and respiratory electron transfer chains
The main idea of the microbial fuel cells and other biohybrid devices is to redirect native electron transfer towards the exogenous acceptors and eventually donate electrons to an electrode. The in vivo organization places adjacent ETC carriers, so the adjacent electron transfer components have appropriate mid-point potentials and a close spatial arrangement.
Two main types of ETCs depend on the activation mechanism of charge transfer: photosynthetic and respiratory. Photosynthetic and respiratory ETCs (PETC and RETC) have different energy sources for electron transfer activation (Fig. 2). In the case of photosynthesis, electron transfer is an endergonic (non-spontaneous) process. The energy of light activates it. Regarding respiration, the energy dynamics of membrane electron transfer are exergonic because the primary donor [NADH (nicotinamide adenine dinucleotide) or succinic acid] possesses a more negative redox potential than the terminal acceptor.28 Different chemotrophic organisms can generate energy in microbial fuel cells.7 Incorporating eukaryotic organisms also provides subcellular compartmentalization, which separates the photosynthetic and respiratory chains into different organelles while simultaneously isolating both chains from the external environment. At the same time, in several prokaryotic organisms, some parts of the ETC can be in the outer membrane. The photosynthetic and respiratory ETCs coexist in the same membrane and have been shown to interact directly with each other.29--31
![[{"id":"9yf2D3LzXN","type":"paragraph","data":{"text":" The fundamental differences between the respiratory and photosynthetic electron transport chains. The respiratory ETC is shown on the left. The photosynthetic ETC is shown on the right. Black arrows indicate electrontransfer pathways. PETC allows both cyclic and linear electron transfer. Instead of oxygen (RETC scheme), other substances can act as terminal electron acceptors in anaerobic respiration. Instead of water (PETC scheme), other substances can act as electron donors in anoxygenic photosynthesis"}}]](/storage/images/resized/gFMt8bO58fxGUzZePxL1LoAXm6I0rJSYmQOkQAcS_xl.webp)
The classification of respiratory electron transport chains is often based on the type of terminal acceptor employed. Aerobic respiration employs oxygen as the terminal acceptor, whereas anaerobic respiration can utilize other compounds such as sulfate and nitrate.32 Furthermore, some microorganisms utilize metal substrates on which they grow as the terminal acceptor for respiration in ETCs. These organisms will be discussed in greater detail below.
The natural diversity of photosynthetic electron transport chains can be easily divided into organisms that can perform oxygenic and anoxygenic photosynthesis. Purple and green photosynthetic bacteria can utilize substances other than water as electron donors and cannot split water; thus, are not able to produce oxygen during photosynthesis. This is often termed anoxygenic photosynthesis. Anoxygenic photosynthesis involves a single type of RC. In contrast, cyanobacteria and plants contain two types of pigment-protein complexes: photosystem 1 (PSI) and photosystem 2 (PSII). These complexes work together to transfer electrons from water to NADP+ (nicotinamide adenine dinucleotide phosphate).
In terms of structure, all ETCs consist of large multi-subunit transmembrane complexes that act as oxidoreductases and lipophilic and hydrophilic mobile electron carriers. The quinone group constitutes the lipophilic carriers in almost all living cells.33 On the other hand, the water-soluble metalloproteins cytochrome c (cyt c) and plastocyanin (Pc) containing heme and copper, respectively, serve as mobile electron carriers. In anaerobic respiration and under iron limitation, flavoproteins also play the role of hydrophilic electron mediators.30
In the case of the PETC, RCs serve as light-induced oxidoreductases.33,34 They produced short-living strong reductants after light quantum energy absorption. Proton gradients across lipid bilayers can be generated by various mechanisms, including the Q-cycle and energy-dependent proton pumps.35,36 The Q-cycle (named for quinol) is a complex mechanism that facilitates the transfer of electrons from quinol molecules to cytochrome complex, generating a proton gradient across the inner mitochondrial or bacterial membrane.37,38
All living organisms contain ETC in their cells. However, only several species are used in the MFC or biohybrid devices:
1) Prokaryotic chemotrophic cells, so-called exoelectrogens, are known as effective organisms for MFC. They include Geobacter sulfurreducens, Rhodoferax ferrireducens and Shewanella sp;39--41
2) Fungi, primarily yeast;42,43
3) Anoxygenic phototrophic bacteria such as purple bacteria. Several works for solar cells are based on the reaction centre isolated from these bacteria;44,45
4) Cyanobacteria can be incorporated into (P)MFC and use their photosynthetic complexes to harvest solar energy for the solar cells;46,47
5) Unicellular eukaryotic algae such as the Chlorella;16,48
6) Vascular plants are the source of isolated photosystems and thylakoid membranes for biohybrid solar cells.49,50
3.1.1. Exoelectrogenic electron transport chains
In nature, there are several unicellular organisms whose chemotrophic metabolisms allow them to reduce exogenous insoluble metal as respiratory terminal acceptors (Fig. 3a). Such organisms are called dissimilatory metal-reducing bacteria (DMRB).51--54 They are commonly used in MFC development primarily because of their ability to generate electricity directly without any modification. Metal-reducing bacteria have unique components on their outer cell membrane that facilitate the transfer of electrons to the outer metal substrate. These components are part of the respiratory electron transport chain and are known as extracellular electron transfer (EET) proteins. EET proteins allow metal-reducing bacteria to use metals as terminal electron acceptors in their respiration process, which is a unique characteristic not found in other types of bacteria. The electrogenic activity of DMRB seems to be provided in two different ways. The first way involves the direct transfer of electrons from extracellular reductases localized in the outer membrane directly to the substrate via membrane multiheme cytochromes. A special case of this path is the transfer of electrons from the cell directly to the electrode along nanowires — special conductive protein filaments.55,56 The second way is mediated by extracellular mobile electron carriers, which leave the cell in a reduced form and donates an electron to the target metal or other acceptor.57 The known, most studied examples of DMRB are Shewanella oneidensis and Geobacter sulfurreducens.
![[{"id":"EShD-8Vpx6","type":"paragraph","data":{"text":"Extracellular electron transfer for DMRB, the central respiratory mechanism (<i>a</i>); yeast, auxiliary respiratory or regulatory mechanisms (<i>b</i>); cyanobacteria (<i>c</i>); algae, auxiliary respiratory and photosynthetic or regulatory mechanisms (<i>d</i>). NADH is not the only candidate for the role of endogenous electron mediator transporting electrons out of the cell. PETC is photosynthetic electron transport chain, PMET is plasma membrane electron transport, RETC is respiratory electron transport chain"}}]](/storage/images/resized/shBbdbMcM1sFmYfi0lOyPxisNX9NuxXZG8LxHA46_xl.webp)
Nanowires discovered in Shewanella are extensions of the outer membrane and periplasmic space.58 The unique metal respiratory system (Mtr) located in these nanowires reduces extracellular metals (Fig. 4). The inner membrane of Shewanella contains NADH dehydrogenase and Q-cycle components, while the outer membrane houses cytochrome transmembrane complexes, which are responsible for transferring electrons to the extracellular acceptor.27 Geobacter nanowires are composed of multi-heme cytochrome proteins such as OmcZ and OmcS that are arranged linearly along the length of the nanowire.59,60 OmcZ is vital for efficient extracellular electron transfer and photocurrent generation in microbial solar cells. OmcS is less conductive than OmcZ but still plays an essential role in EET pathways.61,62 Both Geobacter and Shewanella form conductive biofilm on the metal surface. Biofilms are 3-dimensional structures made up of cells surrounded by self-produced extracellular polymeric substances (EPS) matrix.63 EPS matrix consists of polysaccharides, proteins (cytochromes) and nanowires. Due to the electroactive components present in the EPS matrix, cells can reduce metals even when they are physically separated from the metal surface.
![[{"id":"j-BeL7BbE9","type":"paragraph","data":{"text":" The pathways of direct and mediated extracellular electron transport in DMRB. Three different electron transfer mechanisms are shown from left to right: <i>via</i> a nanowire, <i>via</i> an MTR-like system, and through an extension of the outer membrane and periplasm. IM is inner membrane, OM is outer membrane, cyt is cytochrome, blue arrows indicate electron transfer pathways"}}]](/storage/images/resized/8DYT0AP8VPIIgbDXxVTR1meh59sYpj909x8yapje_xl.webp)
The ability for the electron transfer to the external electrode has been shown for many other heterotrophic bacteria, including E.coli, several acetogenic bacteria, firmicutes, and even archaea.64 However, much less is known about their ETC, as opposed to the ETC of the Geobacter or Shewanella. It also should keep in mind that E.coli has the potential to be used in MFC due to the established set of different metabolic engineering tools.65,66 Many known exoelectrogens, such as the abovementioned Shewanella oneidensis, Geobacter sulfurreducens and Escherichia coli, are proteobacteria.66
3.1.2. Fungi and plasma membrane electron transport
Unlike bacteria, fungi do not have a respiratory chain in their plasma membrane. Instead, respiration in yeasts occurs in mitochondria, and oxygen is the terminal acceptor of the respiratory ETC. Nevertheless, fungi can act as biocatalysts in MFCs due to the presence of ETC in the plasma membrane (Fig. 3b). Plasma membrane electron transport (PMET) is a process that involves the transfer of electrons in the plasma membrane.67 PMET serves as a source of signals derived from changes in the redox state of the microenvironment at the interface of the cytosol and plasma membrane. In fungi, PMET is involved in various metabolic pathways, including fermentation and respiration.67,68 The primary function of PMET in fungi is to maintain the balance between oxidized and reduced NADH. The PMET chain is comprised of the following components: one or more NADH reductase enzymes located on the cytosolic side of the plasma membrane, coenzyme Q within the membrane, hydroquinone oxidases on the outer surface of the plasma membrane that function as terminal oxidases, and an electron acceptor such as oxygen or ferricyanide. PMET is closely related to other metabolic pathways, such as fermentation and respiration.69 PMET was observed in various organisms besides fungi.70--73
In addition to the PMET, some fungi can use an exogenous electron transport system to transfer electrons to extracellular acceptors such as electrodes or other microorganisms due to enzymes such as lactate dehydrogenase and ferricyanide reductase localized in their outer membrane.74
3.1.3. Purple bacteria photosynthetic electron transport chains
The PETCs are particularly interesting because phototrophic organisms can directly produce a photocurrent due to solar energy. Photoheterotrophs, purple bacteria, and some cyanobacteria, can generate current under light and dark conditions due to photosynthetic and respiratory chains. This metabolic versatility increases the applicability of the cells.
The purple bacteria are very metabolically diverse phyla. These bacteria are capable of a range of metabolic pathways. The synthesis of ATP in these organisms can be carried out by aerobic respiration, anaerobic respiration, or anaerobic photosynthesis.45 During aerobic respiration, electrons from NADH or succinate transfer to the molecular oxygen. In the absence of oxygen, some purple bacteria can reduce other organic or inorganic species.75 Sulfides, sulfates, and molecular hydrogen (H2) can all serve as electron donors for the respiration chain in addition to traditional organic compounds such as NADH or succinate.76 The transmembrane protein complexes and respiratory ETC mobile electron carriers are localized in the inner cytoplasmic membrane. In the anaerobic condition, purple bacteria can perform photosynthetic electron transfer in the special intracytoplasmic membranes.77 Oxygen suppresses the synthesis of the photosynthetic pigments of purple bacteria. It leads to the anaerobic character of photosynthesis. The respiratory ETC is linear: electrons are transferred from the primary donor to the terminal acceptor. This electron transfer is conjugated with proton pump operation.
The photosynthetic ETC is predominantly cyclic. The primary donor is the bacteriochlorophyll dimer of the bacterial reaction center. Although there is evidence for external electron transfer in purple bacteria, this process's details and role are unclear.78 The direct electron transfer from purple bacteria to the accepting substrate is believed to be through a mobile quinone pool.45
3.1.4. Oxygenic photosynthetic electron transport chains and cyanobacteria respiration chains
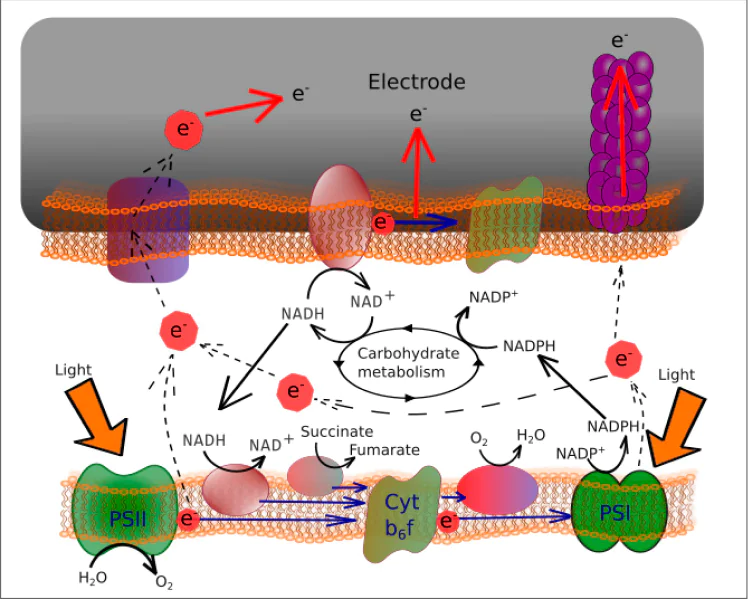
Much attention is paid to studying oxygenic photosynthesis in the context of biohybrid devices. This metabolic pathway needs only light, CO2, and H2O for carbohydrate synthesis. The light reactions of oxygenic photosynthesis results in the synthesis of ATP and NADPH with only an input of light energy and water. This process is followed by the carbohydrate synthesis from CO2 proceeding due to ATP's dephosphorylation and NADPH oxidation. Plants and cyanobacteria have very similar mechanisms of oxygenic photosynthesis with only minor differences (Fig. 5).79 Electrons are transferred from H2O with a redox potential of about +0.93 V to NADPH with a redox potential of –0.4 V. The process requires multiple quanta of light and involves two distinct reaction centres localized within the large pigment--protein complexes PSII and PSI, unlike the single reaction centre in anoxygenic bacteria. The electron transport involves PSI, PSII and b6f complex.28,80 PSII is accountable for the oxidation of water stimulated by light and the reduction of quinone.81 It comprises a unique oxygen-evolving complex that facilitates the process of water oxidation. Cytochrome b6f complex and plastoquinones participate in the Q-cycle.82
![[{"id":"ayIe7P1Bj0","type":"paragraph","data":{"text":" Scheme of oxygenic photosynthesis in plants. FNR is ferredoxin NADP<sup>+</sup> reductase. The Figure was prepared by the authors using original data from the study.<sup>79</sup>"}}]](/storage/images/resized/2KOM9SEcXPVp2puHjH8lX1RSrg4eRwgcM5vhZKUU_xl.webp)
Cyanobacterial photosynthetic apparatus is similar to plants but has several distinctions.1,83 The most prominent feature is that the thylakoid membrane located directly in the cytosol and PETC can interact with respiratory ETC like in purple bacteria. However, photosynthetic ETC is localized solely in the thylakoid membrane, while respiratory ETC is localized both in the thylakoid and inner cytoplasmic membrane. PETC and respiratory ETC share a common plastoquinone pool, b6f complex, and Pc/cyt c pool.31
Cyanobacteria carry out aerobic respiration like mitochondria.84 In such a chain, electrons from NADH (NADPH) or succinate flow to the O2. The respiratory chain contains various dehydrogenases responsible for entering electrons into the chain and a range of terminal reductases.
In addition to photosynthetic and aerobic respiratory ETCs, the outer membrane of cyanobacteria contains ETC components responsible for the electron flow out of the cell (Fig. 3c). The ability of cyanobacteria for extracellular electron transfer has been shown in several research projects.85,86 Gorby et al.55 have shown that Synechocystis PCC 6803 can produce conductive nanowires, like dissimilating metal-reducing bacteria. Cyanobacteria produce nanowires under conditions of CO2 limitation. Moreover, cyanobacteria can transfer electron to the conductive or semiconductive substrate directly and indirectly.87 Direct electron transfer can be facilitated by conductive pili of the cyanobacterial cells.86 This electron excretion mechanism is associated with respiratory and photosynthetic ETCs, which suggests excellent promise for biohybrid devices based on cyanobacteria. While for DMRB (Shewanella oneidensis and Geobacter sulfurreducens), extracellular electron transfer is the primary metabolic pathway (a type of anaerobic respiration), cyanobacteria exploit it as a regulatory mechanism for the export of excess electrons.31 Eukaryotic microalgae can also transfer electrons to an external substrate (Fig. 3d).
4. Microbial fuel cell
A fuel cell is a small portable system that can generate electricity due to the catalytic oxidation of fuel (hydrogen, ethanol, methanol, and other high-energy compounds).88,89 Microbial fuel cell is similar to the traditional fuel cell except that the enzymatic system of the living microorganisms is used instead of specifical metal catalysts.90 It is the device that can generate electricity from biochemical reactions of microbe metabolism.91 MFCs contain two electrodes placed in the same or different vessels (Fig. 6). One of the electrodes, the anode, serves as a terminal acceptor for the extracellular electron transfer of microorganisms immersed in the same vessel. The exoelectrogens oxidize the organic substrate and produce electrons and protons. The other electrode, the cathode, is connected to the anode by an external circuit and an electrolyte connection.90
![[{"id":"WNdoZN0adq","type":"paragraph","data":{"text":" Schematic diagram of microbial fuel cells. Dual-chamber MFC (<i>a</i>); air-cathode single-chamber MFC (<i>b</i>). Blue bold dotted line indicate proton exchange membrane (PEM). Anode was covered by DMRB biofilm in both cases. PEM may not be used in air-cathode MFC"}}]](/storage/images/resized/lHlRbG9FrPwdrWyE0hdCNkGy8cuwd0dRDqkRwhFx_xl.webp)
The use of MFCs is promising in three areas: energy generation, wastewater treatment, and desalination.92 Molecular oxygen serves as a terminal electron sink in many MFCs.93 It plays the same role as in the respiratory process, except that an external electrical circuit is added between the electron transport chain and the terminal oxygen. Microbial electrolysis cells are a particular type of MFC whose primary goal is synthesizing hydrogen gas instead of electricity.17
Numerous designs of MFCs exist, which can be categorized in various ways. We have provided several classification schemes in Table 1.94--118 MFCs with separated anodic and cathodic chambers are called dual-chamber microbial fuel cells (DCMFCs), while single-chamber microbial fuel cells (SCMFCs) have both cathodes and anodes in one container.92,119,120 Fig. 6a shows a classic dual-chamber cell assembled in one vessel. In 2018, Liang et al.121 reported successfully operating a 1000-L modular-designed microbial fuel cell for municipal wastewater treatment. The system consisted of two chambers separated by a proton exchange membrane and could remove up to 90% of chemical oxygen demand from the wastewater while generating electricity.121 SCMFCs are less expensive and easier to assemble but face challenges such as preventing oxygen penetration to the anode.56 One solution is using air-cathodes that reduce oxygen from atmospheric air (Fig. 6b). The presence or absence of a proton exchange membrane is another essential factor in which MFCs differ.92,122,123 Selective exchange of ions between the different chambers is permitted using a proton exchange membrane (PEM) or a salt bridge.122 Polymeric PEM is a more expensive element than a salt bridge.122 At the same time, PEM appears to give an increased efficiency of MFC.124 However, Liu and Logan103 showed that the PEM could be excluded from air-cathode MFCs. Ceramic separators are a less expensive alternative to polymeric PEM.102 In addition, a wide range of distinct electrode materials is used in MFCs.
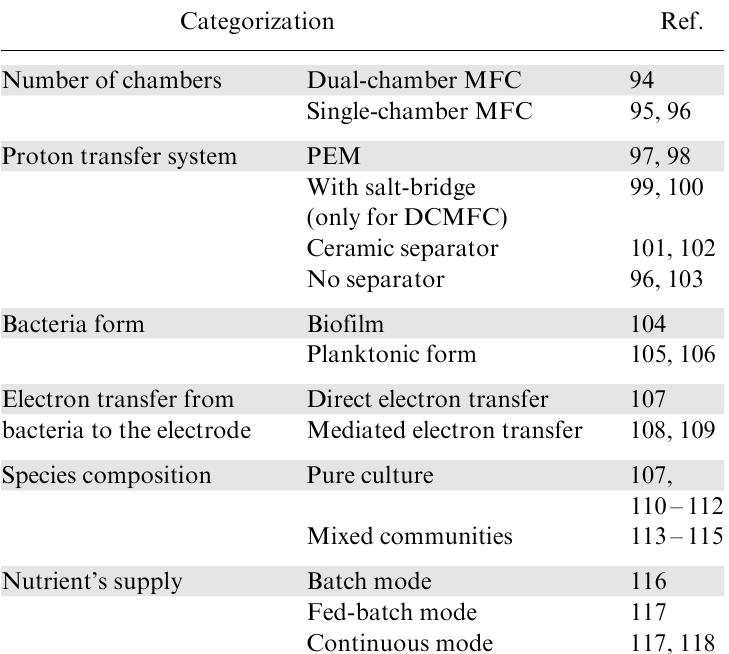
Regarding the microbes used, MFCs can be distinguished by whether they are using pure culture or a mixed community.66 In MFCs, bacteria can exist in two forms: planktonic and biofilm. Planktonic cells are free-floating in the liquid medium, while biofilm cells attach to the electrode surface.93 Biofilms require more time for growth. At the same time, biofilm-based MFC is more stable than the one with planktonic culture.125 However, developing an electroactive biofilm is a significant limitation that must be improved for efficient biocatalysis using a hybrid electrode.63,126
Microfluidic MFCs have been actively researched in the last decade.127 Microfluidic MFCs are small-scale microbial fuel cells that use microchannels and other microfabrication techniques to manipulate fluids at the micrometer scale. Unlike traditional MFCs, which typically use a membrane to separate the anode and cathode compartments, microfluidic MFCs rely on laminar flow to maintain the isolation of independent liquid streams.128
Electrons from bacteria to the anode can migrate in two ways. These ways were outlined in the Section 3.1.1. about exoelectrogenic bacteria. The first way is direct electron transfer (DET), either from the outer membrane protein or from a special nanowire to the electrode. Most often, this is implemented in biofilm-based MFC.112 Another way is the electron transfer between cells and the electrode mediated by a hydrophilic charge carrier. It is called mediated electron transfer (MET). This is realized in MFC with planktonic bacteria and can also be an additional way in biofilm-based MFC. Not all bacteria can emit endogenously produced mediators in the environment. In many studies, an additional exogenous artificial redox shuttle is added to the MFC medium to increase efficiency.66 DET and MET can be realized simultaneously in MFCs.129 However, the efficiency of MFC with DET as the primary electron transport mechanism can be significantly improved by adding an exogenous redox shuttle, thereby increasing the MET activity.108
The success and longevity of a MFC, primarily if intended for bioremediation, depend on its ability to adapt to constantly changing environmental conditions. Research has shown that MFCs containing various consortia of microorganisms are more robust than those with pure cultures.66 Mixed communities are more versatile than single-species cultures. However, the composition of the microorganism community can vary significantly during MFC operation, making it challenging to analyze and achieve stable operation. To address this issue, researchers have developed a co-cultured consortium of microbes as an intermediate option between a wastewater or sludge mixed culture and an axenic culture.130 The members of this community are selected based on their genomics and physiology to create a more stable and predictable system. Co-culture MFCs are a functional upgrade from pure-culture MFCs, with increased stability and performance in several cases.131 A recent study by Liu et al.132 investigated the performance of a consortium-based MFC using a mixture of bacteria-enriched effluent for start-up. The microbial community of the MFC anode biofilm was analyzed using 16S rRNA gene sequence analysis, revealing Geobacter, Geothrix, and Pseudomonas as the predominant bacteria at the genus level. The study found that the MFC with a tungsten carbide decorated carbon cloth (WC/CC) anode exhibited high power density, with a value of 3.26 W m–2 making it one of the best results reported to date.132 However, pure culture MFCs are still highly interesting as analytical tools for investigating external electron transfer.57,66 They can also be used for comparative analysis of the activity of various mutants with wild-type bacteria in the same system, which is another method for studying bacterial metabolism and increasing the productivity of MFCs.66
MFCs can be designed in three modes: batch, fed-batch, and continuous modes.133 In batch mode, nutrients, and growth medium are added together within the starting volume at the start of the bioprocess. Although easy to set up, batch mode MPCs are less efficient due to an initial log phase of microorganism growth. They can also suffer from the accumulation of toxic metabolites over time.134 The fed-batch mode of MPC allows selected nutrients to be added in stages throughout the bioprocess when electrical output falls below a baseline. Although this offers more control of the MFC performance, it also increases the possibility of introducing contamination and can still suffer from the accumulation of toxins.133 The fed-batch approach can more rapidly reach and maintain the exponential growth phase. Continuous mode involves a constant flow of nutrients into the MFC, which allows for stable operation but requires more complex equipment and maintenance. In this mode, a new culture is added while microalgal biomass is removed simultaneously, avoiding toxins accumulation.116 This can reach a functional plateau and avoid the accumulation of toxins.
The performance of MFCs depends on various factors, including the substrate, electrode material, inoculant composition, and MFC construction features mentioned above. The substrate is the primary energy source in MFCs and can be pure compounds like acetate, or sucrose, or organic matter from wastewater for remedial functions.135,136 Electrode modifications are an essential strategy for improving microbial fuel cell performance and increasing their potential applications in various fields, such as wastewater treatment and renewable energy production. Frмhauf et al.137 distinguish two significant strategies for electrode surface modifications: increasing available surface area and altering electrode surface chemistry. Their study explores the electrostatic self-assembly of charged polymers to modify the surface of indium tin-coated electrodes and improve microbial fuel cell performance. Modifying electrode surfaces can increase the available surface area for cells to adhere to, increasing current production. Additionally, altering the surface chemistry of electrodes can influence cellular adhesion, affecting the start-up time and initial phase of biofilm formation. Carbon materials, copper, stainless steel, indium tin oxide, titanium, platinum, and gold are commonly used for anodes in MFCs.56,90,133,138,139 Suitable species for MFCs include acidobacteria, firmicutes, proteobacteria, yeasts, and cyanobacteria.17
The majority of bacteria studied in MFCs, including DMRB, are Gram-negative bacteria. Unlike Gram-positive bacteria, these microorganisms lack a thick cell wall that separates the ETC membrane from the electrode. Gram-positive bacteria have a thick layer of peptidoglycan that separates their cell membrane from the external environment. Due to this structural difference, Gram-positive bacteria are considered less efficient electricity generators in MFCs. Consequently, possible mechanisms for electron transfer to an external acceptor in Gram-positive bacteria are less studied than in their Gram-negative counterparts. Pankratova et al.140 conducted a study on the Gram-positive lactic acid bacterium Enterococcus faecalis, in which they demonstrated the bacterium's ability to transfer electrons generated in fermentation metabolism to electrodes both directly and indirectly via mediators. An osmium redox polymer was used on the graphite electrode to encapsulate bacterial cells. Interestingly, their findings showed that heme proteins are not required for EET, which contrasts with the EET of DMRB. The study also concluded that quinone is essential for the EET of E.faecalis. It appears that the positively charged osmium redox polymer network on the electrode may intercalate with the abundant negatively charged teichoic acids in the cell envelope, allowing it to reach positions close enough to the membrane to facilitate efficient electron transfer from reduced quinones to the electrode.
Unicellular fungi, primarily yeast, which also possess a cell wall, are more commonly utilized as biocatalysts in MFCs compared to Gram-positive bacteria. However, their implementation in MFCs is still considerably less researched than that of Gram-negative bacteria. Yeast is the most used type of fungi in MFC research due to its ability to use a broader range of organic compounds as a source of energy, including complex compounds such as starch and cellulose. Yeast can also thrive under anaerobic conditions, making them more resistant to environmental changes. Additionally, yeast production is relatively simple and does not require complex conditions.141
Fungi can act as catalysts in the cathode chamber of MFCs and play an essential role in the electron transfer system of bacterial-based microbial fuel cells. For example, Trametes versicolor and Ganoderma lucidum can produce laccase enzymes that catalyze oxygen reduction at the cathode chamber.142,143 In addition to using single microorganisms as catalysts in MFCs, some studies have investigated the use of mixed cultures of bacteria and fungi. These mixed cultures can take advantage of the complementary metabolic pathways of different microorganisms to improve MFC performance. Fernвndez de Dios et al.144 used a mixed culture of S.oneidensis MR-1 and Trametes versicolor to achieve higher power densities than either microorganism alone. The maximum volumetric power density achieved was 1.5 W m–3.
In Table 2, a summary of the performance of various MFCs is presented based on articles published within the last three years.42,132,145–152
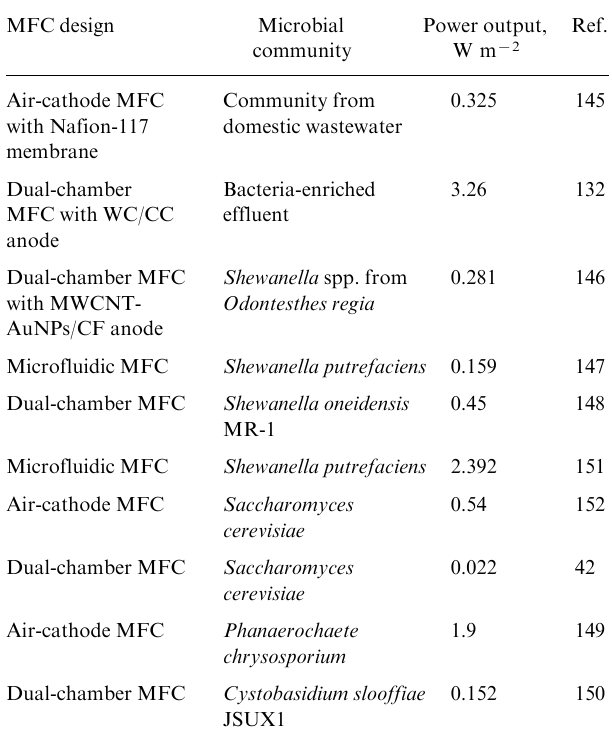
Notably, Rewatkar and Goel151 achieved the best results for pure culture MFCs in 2022, using Shewanella putrefaciens to power a microfluidic MFC. The microfluidic device featured a Y-shaped microchannel made of polydimethylsiloxane and printed circuit board bioelectrodes. The researchers used photolithography techniques to fabricate the printed circuit board bioelectrodes and patterned them with multi-walled carbon nanotubes (MWCNTs) to enhance their performance. The Y-shaped microchannel design separated anolyte (S.putrefaciens) and catholyte (potassium ferricyanide) from the same microfluidic channel, creating a virtual membrane. The co-laminar fluid flow behaviour of the device facilitated the maintenance of the separation between the two fluids. Through optimizing bacterial volume and flow rate conditions, the researchers achieved a maximum power density of 239.2 μW cm–2, with a bacterial volume of 10 μL and a flow rate of 0.5 μL min–1 being the optimal conditions. In the field of fungal based MFC, Sukri et al.149 achieved the best performance. Their single-chamber, membrane-less enclosure utilized P.chrysosporium (a fungi derived from white-rot) fed with agro-waste substrates and paired with a zinc anode. In contrast to most MFCs, the fungal microbes were not cultured on the current collector but were left to be freely suspended in the unbuffered electrolyte. This straightforward and self-sufficient design could continuously sustain a 1 mA discharge current for 44 days, with an average operating voltage of 0.67 V (1056 mA h capacity). During continuous operation, this MFC achieved a maximum power density of 1.9 W m–2 with an open-circuit voltage of 0.8 V and a short-circuit current density of 3.5 A m–2. These results demonstrate the potential of fungi-based MFCs for sustainable and efficient power generation using waste materials as a substrate. The best results among all MFCs are shown by fuel cells based on microbial communities.132 The three-electrode configuration is commonly used for investigating bioelectrodes, and the two-electrode MFCs.145,153,154
5. Photosynthetic microbial fuel cell
PMFC is similar to the MFC. They differ by primary energy source for the living bacteria (Fig. 7a). Phototrophic microorganisms in PMFC obtain energy from the light, while bacteria in MFC obtain energy from the chemical substrate.155 Many PMFCs can convert sunlight and chemical bond energy of organic substrate into electricity due to the flexible metabolism of photosynthetic bacteria. PMFCs can be classified in the same ways as MFCs.
![[{"id":"7GeZuDFAQV","type":"paragraph","data":{"text":"Schematic diagram of a photosynthetic microbial fuel cell. Dual-chamber PMFC (<i>a</i>); three electrode configuration in single-chamber PMFC (<i>b</i>); algal-assisted PMFC (<i>c</i>). Subs is substrate, CE is counter electrode, RE is reference electrode, WE is working electrode"}}]](/storage/images/resized/YnmxFDpVvFpPsajIU2NZYbPIKkp0DNXOQmRDBqF4_xl.webp)
For practical energy conversion applications, a PMFC with two electrodes should be both cost-effective and efficient and should not necessitate an external power source. However, in the case of studying the interaction between microorganisms and electrodes, the three-electrode scheme is indispensable (Fig. 7b). The three-electrode scheme makes it possible to obtain a current-voltage diagram and evaluate the energy levels of the electrodes.156 Also, the three-electrode scheme makes it possible to carry out chronoamperometric measurements at various applied voltage biases. In this chapter, we will consider electrochemical cells with both two and three electrodes. The efficiency of PMFC includes:
1) the absorption capacity of photosynthetic pigments;
2) the quantum yield of the photosynthetic reaction centre;
3) the efficiency of electron transfer to the electrode;
4) the efficiency of the reduction reaction at the cathode.
This paragraph focuses on the interaction of the bacterial cell with the electrode.
Cyanobacteria are often utilized in biohybrid photodevices due to their diverse electron transport pathways and easy adaptation to changing environmental conditions.31,157 Due to this, most of the described PMFCs contain cyanobacteria instead of anoxygenic phototrophs. Fewer publications are associated with eukaryotic algae (Chlamydomonas, Chlorella).156,158--160 The thylakoid membrane in the green algae is localized in the chloroplast. An additional membrane between the thylakoids and the electrode seems to contribute to the resistance and prevent photocurrent generation. Another advantage of cyanobacteria over green algae is the ease of genetic manipulation.161 Anoxygenic purple bacteria are also of great interest for the development of PMFCs.45,78,162 Many studies are devoted to PMFC based on a mixed consortium of microorganisms, including MFC with phototrophic and chemotrophic organisms.46,110
The pioneering research on the interaction between photosynthetic organisms and electrodes was performed in the early 1960s. Berk and Canfield163 built the PMFC containing Rhodospirillum rubrum and cyanobacterial community in the anodic and cathodic chambers, respectively. R.rubrum generated hydrogen under light, and cyanobacteria produced oxygen under the light. On the platinum electrodes, hydrogen oxidized, and oxygen reduced in the anodic and cathodic chambers. In the cell with platinum electrodes, the open-circuit potential was 0.917 V, and the short-circuit current density was 162 μA cm–2. Despite the notable result, the authors expressed doubt that such devices would outperform conventional fuel cells in terms of efficiency.
The PMFCs began to actively study in the 80s of the twentieth century.164--166 The investigation of the electrical signal from the chloroplast, thylakoids, and isolated reaction centers preceded the PMFC study.24,167 In 1979, Ochiai et al.168 reported a big anodic photocurrent upon the three-electrode cell with chloroplast-contained electrode. The authors used broken chloroplasts from spinach without or with minor remnants of the outer membrane or stroma. There were so-called "type C" chloroplasts following the obsolete classification of D.O.Hall.169 These membrane preparations were entrapped in the polyvinyl alcohol matrix, followed by deposition onto the SnO2 optically transparent electrode. The photocurrent in this system is as much as 1 μA cm–2 per 10 μg chlorophyll under illumination 60 lux at the bias of 0.5V vs. saturated calomel electrode (SCE). At the same time, authors meet problems of stability and longevity of such bio-based photoconverters due to the high susceptibility of the isolated chloroplasts. For these reasons, the authors switched to photoconverters based on thermophilic cyanobacteria Mastigocladus laminosus, which usually inhabit hot springs.164,168 The living algae electrode was obtained by depositing the microalgae immobilization in the calcium alginate matrix onto the SnO2 optically transparent electrode. The immobilized M.laminosus cells retain the ability to grow. During the 20 days of continuous illumination, an electrochemical system with a cyanobacterial electrode shows increasing accumulated chlorophyll and photocurrent. The photocurrent attained 40 nA cm–2 at illumination adjusted to 50 kilolux. There are no electron shuttles in this system. A Pt plate and an SCE are the counter and reference electrodes respectively. All three electrodes were immersed in the same vessel. It is important to note that this system should not be referred to as a PMFC since the photocurrent signal was only recorded when a bias voltage was applied. Nevertheless, this pioneering work initiated the study of PMFCs. In a follow-up study, Ochiai et al.165 demonstrated that the addition of the photosynthesis inhibitor 3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU) decreased the photocurrent output and exogenous electron acceptor (mediator) increased it.
Subsequently, the researchers of biological photocurrent moved away from thermophilic bacteria. Anabaena, Synechococcus, and Synechocystis species are most often used in the research.166,170,171 Perhaps this is due to more accessible cultivation methods for mesophilic cyanobacteria. Later, bacteria such as Nostoc, Leptolyngbya, Chroococcales, and Lyngbya were used.157,172,173 The first PMFC, to the best of our knowledge, was built by Tanaka et al.166 and described in 1985. The design of their fuel cell was similar to the conventional MFCs actively researched during those years.174,175 It was a dual-chamber fuel cell constructed from polymethacrylate plastic with a carbon anode and a platinum foil cathode. Anabaena variabilis strain M-2 was inoculated in the anodic chamber. They used 2-hydroxy-1,4-naphthoquinone (HNQ) as an effective electron mediator in the anodic chamber instead of thionine. Thionine was an effective electron mediator in MFC based on the chemotrophs. However, its exploitation in PMFC failed because of its instability under the light. The cathode solution contained ferricyanide as an electron acceptor from the cathode. The current in this cell attained 1-2 mA immediately after running and then decreased for several hours. Also, it was shown that old Anabaena cells (200 h of cultivation) were more effective in the PMFC than young cells (100 hours of cultivation). There was no exogenous substrate, but the older cell culture accumulated more endogenous glycogen than the younger culture.
It is worth noting that MFCs based on the mixotrophs can generate current both under light and dark due to the photosynthesis and oxidation of the substrate, respectively. Depending on the type of bacteria, their metabolism, and the number of accumulated carbohydrates, the dark current may be higher or lower than the photocurrent. A team of researchers from the National Institute of Resources and the Environment of Japan investigated the relationship between dark and light currents in MFCs based on cyanobacteria.171,176--179 In 1993, Yagishita et al.170 demonstrated that the inhibition of the photosynthesis by carbonyl cyanide m-chlorophenylhydrazone (CCCP), DCMU, 2,5-dibromomethylisopropyl-p-benzoquinone, phenylmercury acetate and N,Nʹ-dicyclohexylcarbodiimide lead to the decrement of the current generation. They used Synechococcus sp. and HNQ in the anodic chamber. In a subsequent study, Yagishita et al.179 constructed a PMFC utilizing A.variabilis M-3 immobilized within alginate beads. The primary current source for this cell was the result of carbohydrate oxidation. The PMFC was discharged in the dark after the anode and cathode were connected through an external circuit. The authors increased the functional lifespan of this fuel cell by alternating periods of discharging in the dark with periods of accumulation of hydrocarbons in the light. In 1999, the same group177 analyzed exogenous glucose addition impact on the PMFC activity. They exploited Synechocystis sp. PCC6714. This strain has a higher chemoheterotrophic activity than other cyanobacteria. The authors showed that the more exogenous substrate added, the greater the current will be generated both under illumination and in the dark. In this investigation, the dark current was higher than the light current. Sparging the anode chamber with nitrogen gas provides anaerobic conditions and promotes current generation during illumination.171 Yagishita et al.178 selected the right light conditions, Synechococcus cell density, and HNQ concentration, and the energy conversion efficiency was 3.3% in their PMFC. However, at higher cell densities, current generation was suppressed by light scattering; at a higher concentration of HNQ, the excess of oxidized HNQ absorbed a significant part of the photosynthetically active radiation, which also led to a decrease in photocurrent; at a higher light intensity, HNQ was oxidized by excess photosynthetically generated oxygen, i.e., a short circuit occurred in the anode chamber, and cell efficiency decreased. This PMFC generated a maximum power density of about 0.2 mW cm–2. The following reaction occurred in the anodic chamber:
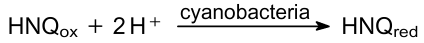
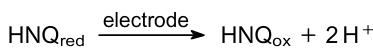
HNQ was a popular mediator for PMFC in earlier works. It can accept electrons from fumarate and nitrate reductase regulator (FNR) and NAD(P)H dehydrogenase in the case of light and dark conditions, respectively.178 However, the reduced form of HNQ can reduce oxygen and, in such a way, shorten circuit. Due to this, HNQ is the mediator of choice in the case of anaerobic conditions for heterotrophic mod of cyanobacterial MFC.177 Early research on characterizing suitable electron mediators was performed by Martens and Hall,180 Tsujimura et al.,181 and Torimura et al.182
Martens and Hall180 also drew attention to the hydrophobicity and toxicity of exogenous mediators. They compared three mediators: diaminodurene (DAD), ferrocyanide, and p-benzoquinone (BQ). Synechococcus cells immobilized in a polymer matrix deposited on the electrode were used. A three-electrode setup was used to study the electrodes. The effectiveness of DAD was higher than that of ferrocyanide and p-benzoquinone. Ferricyanide is a poor choice as an electron acceptor for whole cyanobacterial cells due to its hydrophilicity and inability to penetrate the cytoplasmic membrane. In addition, the authors hypothesized that p-benzoquinone inhibits PMFC activity upon long-term exposure. Another characteristic of the mediator is the place of electron acceptance in the electron transport chain. For clarity, it is worth mentioning that in the more recent studies, ferricyanide successively mediated an electron transfer between the planktonic Synechocystis culture and indium-tin-oxide anode.183 It was suggested that ferricyanide obtains electrons from the putative trans-membrane protein at the cell surface.
Tsujimura et al.181 used the 2,6-dimethyl-1,4-benzoquinone (DMBQ) as an electron acceptor in the PMFC anodic chamber with the Synechococcus sp. PCC7942 cells.
The reactions in the anodic chamber include electron transfer from ETC to the DMBQ:
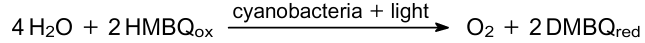
And electrode reaction of DMBQ:
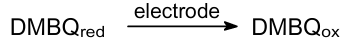
The researchers performed both PMFC performance measurement and characterization of half-cell reactions in the cathode and anode chambers separately. The PMFC generated a maximum power density of about 0.03 mW cm–2 (Ref. 181). The authors showed that the activity of the anodic mediator depends on its redox potential, permeability through the cyanobacterial membrane, and the kinetics of the electrode reaction. Torimura et al.,182 with the rotating disk electrode technique, compared the acceptor ability of two quinones: BQ and DMBQ. They have shown the kinetics of the quinone reduction by Synechococcus sp. PPC7942 cells under illumination behave as a Michaelis-Menten reaction type for the mediator concentration and the light intensity. DMBQ seems superior to BQ for photoreduction. They used Synechococcus sp. cell-entrapped and DMBQ-embedded carbon paste electrodes. Different mediators accept electrons from different sites of the ETC. BQ and DMBQ accept electrons from the PSII acceptor side, HNQ accepts electrons from the PSI acceptor side, and ferricyanide accepts electrons from the PSI acceptor side. Shlosberg et al.184--186 found that NADPH can act as an efficient electron mediator between PSI of the cyanobacteria or microalgae and the graphite anode, leading to enhanced photocurrent generation.
Developing PMFCs with mediators in the anode compartment as scalable devices is challenging due to several factors. Firstly, using mediators add to the overall cost of the system. Secondly, the efficiency of PMFCs can be negatively impacted by the deterioration of mediators, which can lead to issues such as toxicity and short-circuiting.87 A suitable alternative to dissolved mediators is a conductive polymer layer on the anode in which cells are fixed. Such a layer performs both the function of immobilization and the function of electron transfer. Furukawa et al.187 reported about solar cells without mediators. In that work, PMFC with polyaniline treated carbon cloth electrode mediator was compared with a similar cell with the untreated electrode. The anode chamber of the first cell did not contain mediators; the second cell contained methylene blue as a mediator. They used Synechococcus. sp in the anodic chamber. The maximum power of the cell with polyaniline was several times greater than that of the cell with the mediator. Zou et al.46 constructed a single-chamber PMFC that did not depend on an organic substrate. The anode of this PMFC contained conductive polymers. More recently, this group modified PMFC anodes with a nanostructured electrically conductive polymer, polypyrrole.188 In this research growing fresh-water photosynthetic biofilm was used.
In 2014, Sekar et al.157 wired the Nostoc sp. ATCC 27893 to carbon nanotubes (CNTs). CNT also serves as an extracellular electron acceptor. The PMFC utilizing an exogenous mediator generated a maximum current density of 250 mA m–2 (25 μA cm–2) and a peak power density of 35 mW m–2. CNT function similarly to a conductive polymer matrix. CNTs fix cells and accept electrons. McCormic et al.189 built PMFC with a biofilm of the pure microalgae culture that produces a photocurrent without any mediator or polymer layer. The green alga or cyanobacteria cells were grown directly on an indium tin oxide-coated polyethylene terephthalate electrode. Cyanobacteria have shown greater electrogenic capacity than green algae. A series of PMFCs generated enough power to run a digital clock. Biofilm on the anode can realize DET due to an electron-transfer enzyme in the outer membrane or an exogenous electron mediator. In the first case, only a single layer of cells can be used for electricity generation. Saper et al.190 constructed PMFC with Synechocystis sp. PCC 6803 cells applied by gravity onto the graphite electrode. In this PMFC, the current was generated due to endogenous electron mediator. However, mild cell pretreatment is required to increase efficiency. The microfluidizer was used for this pretreatment. The photocurrent was about 35 μA cm–2 at the applied potential of 150 mV vs. Ag/AgCl. For all their shortcomings, mediators significantly increase the photocurrent.157
Tsujimura et al.181 pioneered using a catalytic system in a cathode chamber for oxygen reduction. In previous studies, the cathode chambers contained excess potassium ferricyanide, which served as a sacrificial electron acceptor.103,191 The practical exploitation of cells with a sacrificial component is difficult and ultimately limited due to the depletion of mediators. The authors proposed to use bilirubin oxidase (BOD) acting in tandem with 2,29-azinobis(3-ethylbenzothiazolin-6-sulfonate) (ABTS2-) in the cathodic chamber. BOD catalyzes the reaction:
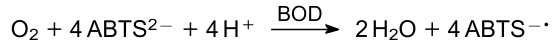
Oxidized ABTS gains electrons again at the cathode:

Sekar et al.157 made a similar scheme with laccase enzyme instead of BOD. It has been shown that multicopper laccase oxidase supported on carbon nanotubes can catalyze the bioelectrochemical reduction of oxygen. This system does not require any mediator compared to soluble BOD/ABTS2–.
Electron transfer efficiency from the ETC to the anode is determined by choice of organism and mediator and the anode material. It must be stable, biocompatible, and conductive. The anode should be transparent. Using an opaque anode imposes additional restrictions on the design of the entire cell. The most used electrodes are carbon materials (foil, felt, paper, etc.), metal plates (gold, platinum), and, more recently, glass electrodes with a thin layer of transparent conductive oxide (indium tin oxide and fluorine-doped tin oxide).93
Bioelectrochemical cells based on the components of the photosynthetic apparatus (thylakoid membrane, PSI, PSII complexes, bacterial reaction centres) are investigated in parallel with PMFC based on the whole microorganisms. Bombelli et al.183 fabricated a photovoltaic device that allowed direct comparison between the whole bacterial cell and isolated photosynthetic complexes. This PMFC functioned without bubbling and external voltage bias. They compared Synechocystis sp. PCC6803 cells and thylakoid membranes from Spinacia oleracea in their device. The thylakoids have demonstrated a higher efficiency in converting light into electricity. However, cyanobacteria could generate current both in the dark and in the light, and this generation was more stable than in the case of isolated membranes.
Several reports of mutant strains of PMFC have been published in the last decade. Measurement of mutant strains' photocurrent and/or dark current can serve two purposes.159,192,193 The first goal is the study of mutants. In this case, comparing the photocurrent signals of the wild type and the mutant strain makes it possible to evaluate the effect of the mutation on photosynthesis and extracellular electron transfer in bacteria or algae. The second goal is the selection of strains for the most efficient functioning of PMFC. Bradley et al.194 analyzed the electrogenic activity of the cyanobacterium Synechocystis sp. PCC 6803 mutants without terminal respiratory oxidases. The removal of various respiratory terminal oxidases has been shown to increase power output. This result proved that PSI mediates electron transfer from the substrate to the electrode the in the investigated system. Longatte et al.160 has focused on the PSII state of the Chlamydomonas reinhardtii in the PMFC with different quinone-type exogenous mediators. Due to minimizing the PSI involvement in the PETC, they used a mutant, ΔpetA, that lacks the b6f complexes. Kusama et al.192 has shown that the outer membrane of Synechocystis sp. PCC 6803 limits the extracellular electron transfer and photocurrent. They compared a mutant strain of Synechocystis that lacks the outer membrane, slr0688i, with wild-type, dCas9, in a single-chamber PMFC with an ITO-coated glass anode, a Pt cathode, and an Ag/AgCl reference electrode.
Lewis et al.193 investigated the ability of Synechocystis ΔpsbB mutant cells lacking PSII to generate cathodic photocurrent in the presence of duroquinone (DQ) as an exogenous electron mediator. The reason for the removal of PSII is the low stability of PSII and increased generation of reactive oxygen species in PSII in the presence of light. Both electrodes were carbon clothes. The cathodic chamber also contained Ag/AgCl reference electrode. The cathode was held at a constant potential of –0.249 V to facilitate DQ recovery. The PMFC yielded an average current density of 150 A m–2 s–1 mg-chl–1 under light intensity up to 2050 μmol photons m–2 s–1. It was suggested that electrons from reduced DQ go to the PSI through the b6f complex. The cyclic pathway around PSI also functions in this system. Adding a photosynthesis inhibitor is another way to study electron transfer between the ETC and the electrode in PMFC. Inhibitor impact depends differently on the PMFC design and cyanobacterial specie. This way, photocurrent from Nostoc and Lyngbya was reduced by DCMU and CCCP; however, the addition of DCMU to the Synechocystis suspension increased the photocurrent.172,190 Both of the systems mentioned above generated photocurrent without any exogenous mediator. Table 3 summarizes the performances of PMFC reported in the last three years.193,195--198

Microalgae have recently emerged as promising photocatalysts in developing of algal-assisted microbial fuel cells (AAMFCs).199 These DCMFCs employ microalgae or cyanobacteria in the cathode chamber to generate bioelectricity. Since algae are photosynthetic organisms that can produce chemical energy through photosynthesis with solar energy, live algae in the cathode chamber can act as a catalyst for oxygen reduction reaction, thereby enhancing the performance of the MFC (Fig. 7c). Alternatively, dead algal biomass or algal powder can be used in the anode chamber as a substrate for heterotrophic microbes. This unique feature of the AAMFC allows for simultaneous wastewater remediation, algal biomass recovery, and bioelectricity production, thereby making it a promising approach for wastewater treatment. Table 4 summarizes the performances of various AAMFCs reported in the last three years.200--203

6. Conclusion
Over the past 30 years, there have been significant advancements in the study of PMFCs. However, the electrical signal from PMFCs is still lower than that of MFCs based on chemotrophic microorganisms.192 Additionally, extracellular electron transfer of photosynthetic bacteria remains a significant fundamental problem. Nonetheless, studying PMFCs contributes to the solution to this problem. It is unlikely that pure culture-based PMFCs will have practical applications due to low yield and high maintenance costs. PMFCs based on mixed cultures are more promising.158 Fuel cells that combine photo- and chemotrophic organisms are also fascinating. Fuel cells that use the microflora of the rhizosphere or the microflora of bottom sediments in combination with photosynthesis are particularly noteworthy.204 Despite their low efficiency, such cells could occupy a niche in the energy converter market if their cost and maintenance costs were minimized.
No standard conditions for testing PMFC or MFC make it difficult to compare individual cells. However, several reviews attempt to compare the results of different studies conducted by different groups, providing insight into the current situation in the PMFC field.17,93,126,205
This work was supported by the Russian Science Foundation (Project No. 19-14-00118). Fig. 2 was supported by a state contract of the Ministry of Science and Higher Education of the Russian Federation (Project No. 122050400128-1). BDB was supported by the Charles P. Postelle Distinguished Professorship at the University of Tennessee.
7. List of abbreviations
AAMFC --- algal-assisted microbial fuel cell,
ABTS --- 2,29-azinobis(3-ethylbenzothiazolin-6-sulfonate),
ADP --- adenosine diphosphate,
ATP --- adenosine triphosphate,
BOD --- bilirubin oxidase,
BQ --- 1,4-benzoquinone,
CCCP --- carbonyl cyanide m-chlorophenylhydrazone,
CE --- counter electrode,
CNT --- carbon nanotube,
cyt --- cytochrome,
DAD --- diaminodurene,
DCMFC --- dual-chamber microbial fuel cell,
DCMU --- 3-(3,4-dichlorophenyl)-1,1-dimethylurea,
DET --- direct electron transfer,
DMBQ --- 2,6-dimethyl-1,4-benzoquinone,
DMRB --- dissimilatory metal-reducing bacteria,
DQ --- duroquinone,
EET --- extracellular electron transfer,
EPS --- extracellular polymeric substances,
ETC --- electron transport chain,
HNQ --- 2-hydroxy-1,4-naphthoquinone,
IM --- inner membrane,
MET --- mediated electron transfer,
MFC --- microbial fuel cell,
Mtr --- metal respiratory system,
MWCNT --- multi-walled carbon nanotubes,
NADH/NAD+ --- nicotinamide adenine dinucleotide,
NADPH/NADP+ --- nicotinamide adenine dinucleotide phosphate,
OM --- outer membrane,
Pc --- plastocyanin,
PEM --- proton exchange membrane,
PETC --- photosynthetic electron transport chain,
PMET --- plasma membrane electron transport,
PMFC --- photosynthetic microbial fuel cell,
PSI --- photosystem I,
PSII --- photosystem II,
RC --- reaction centre,
RE --- reference electrode,
RETC --- respiratory electron transport chain,
SCE --- saturated calomel electrode,
SCMFC --- single-chamber microbial fuel cell,
WC/CC --- carbide decorated carbon cloth,
WE --- working electrode.