Keywords
Abstract
The research into quorum sensing inhibitors is mainly directed towards fighting pathogenic bacteria resistant to therapy with antimicrobial drugs. Natural quorum sensing inhibitors may serve as an effective alternative to antibiotics, since they are less likely to cause the development of drug resistance of pathogenic microbiota. Currently, the search for effective inhibitor molecules and analysis of structure–activity relationships among natural compounds is considered to be the most obvious strategy towards solution of the antibiotic resistance problem. Since plants have co-evolved with the microbial environment and have been constantly exposed to bacterial infections, they successfully developed chemical mechanisms to fight external pathogens, in particular the quorum sensing inhibition. The review considers the secondary metabolites of various classes of plants such as terpenes, quinones, coumarins, stilbenes, alkaloids, curcuminoids, flavonoids, phenolic compounds, and their derivatives and analogues that possess quorum quenching activity. Some aspects of the structure–activity relationships for the described compounds are highlighted for the first time.
The bibliography includes 139 references.
1. Introduction
Antimicrobial resistance is a serious socioeconomic problem recognized in many developed countries as a threat to national security. Infectious diseases caused by drug-resistant microorganisms are protracted; they more often require hospital admission of patients and longer time spent in the hospital and also deteriorate the prognosis for full recovery. In those cases where the drugs of choice (first-line drugs) do not have the desired effect, it is necessary to use second- and third-line drugs, which are often more expensive, less safe, and have limited availability. All these factors increase the direct and indirect financial costs of treating infections and increase the probability of spread of resistant microorganisms in the population.[1]
It follows from published data that ~ 90 – 95% of strains of Staphylococcus aureus all over the world are resistant to penicillin, and in most of Asian countries, the percentage of methicillin-resistant strains of Staphylococcus aureus (MRSA) is 70 – 80%. The MRSA-caused infections are treated with vancomycin, but vancomycin-resistant S. aureus (VRSA) strains have also already appeared.[2] A similar situation is observed for Pseudomonas aeruginosa: multiresistant or even panresistant strains of this Gram-negative bacterium have been reported.[3][4] Therefore, a highly relevant task is to search for new approaches to control infections, including those caused by drug-resistant strains, without using antimicrobial agents or using lower doses of antimicrobial agents. Bacterial quorum sensing (QS) inhibitors may become a promising alternative to antimicrobial drugs.
Quorum sensing is an established biological term meaning cell-to-cell communication widely used by bacterial pathogens to coordinate the expression of several collective traits, including the production of virulence factors, biofilm formation, and swarming and other types of motility as the population threshold has been reached. There are numerous facts indicating that QS enhances the virulence of bacterial pathogens in animal models and in human infections.[5][6] This communication is accomplished by signalling molecules called autoinducers (AIs).[7]
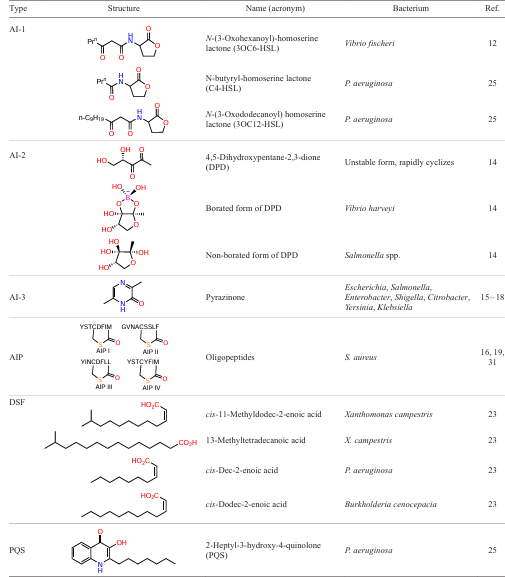
In Gram-negative microorganisms (including Aeromonas, Brucella, Burkholderia, Pseudomonas, Serratia, and Yersinia pathogenic bacteria),[8] a major group of AIs are N-acyl homoserine lactones (AHLs), which are also called AI-1.[6][9] Gram-negative bacteria use these compounds, first of all, for intraspecies communication,[10] although some publications indicate[8][11] that AHLs and their analogues also act on QS systems of S. aureus strains. All currently known AI-1 contain a homoserine lactone ring with a fatty acid residue at the nitrogen atom (in position 1). Acyl groups vary in chain length (from 4 to 18 carbon atoms), saturation levels, and oxidation states, and in most cases, the chain consists of an even number of carbon atoms.8 These differences determine the specificity of AHLs.[12]
The first studied QS system of Gram-negative bacteria is the LuxR/LuxI system of the bioluminescent bacterium Vibrio fischeri mediated by the N-(3-oxohexanoyl)-homoserine lactone (3OC6-HSL) autoinducer. The QS systems of most other Gram-negative bacteria are homologous to this system. The QS system of P. aeruginosa consisting of LasI/LasR and RhlI/RhlR systems is regulated by two autoinducers, N-(3-oxododecanoyl)- (3OC12-HSL) and N-butyryl- (C4-HSL) homoserine lactones, respectively. These systems regulate the expression of genes important for bacterial virulence (e.g., elastase and alkaline phosphatase genes and genes involved in the synthesis of the pyocyanin pigment) and the synthesis of virulence factors (elastase, protease, and alkaline phosphatase enzymes and exotoxin).[10][12]N-Acyl-homoserine lactone-mediated QS systems are among the best studied ones; therefore, the search for molecules that effectively inhibit this process among structural analogues of these systems is presumably a promising research strategy.[13]
As opposed to AI-1, which, as noted above, are mainly used by bacteria for intraspecies communication, a different type autoinducer (AI-2) is also involved in interspecies communication of both Gram-negative and Gram-positive bacteria. Structurally, AI-2 is unstable 4,5-dihydroxypentane-2,3-dione (DPD), which spontaneously cyclizes to borated (e.g., for Vibrio harveyi) or non-borated form (e.g., for Escherichia coli or Salmonella spp.). Depending on the type of bacteria, AI-2 participates in the regulation of quite a few functions: bioluminescence, biofilm formation, production of virulence factors, susceptibility to antibiotics, motility, chemotaxis, cell adhesion, growth in iron-limited medium, etc.[14]
Another interspecies QS communication system is the AI-3/adrenaline/norepinephrine system used by the Escherichia, Salmonella, Enterobacter, Shigella, Citrobacter, Yersinia, and Klebsiella bacteria.[15][16] Currently, this system has not been comprehensively studied as yet, since the exact chemical structure of AI-3 is obscure. However, it is known that at least some AI-3 are pyrazinones[17][18] and that AI-3-mediated systems regulate flagella genes and pathogenicity as well as participate in signal transduction between kingdoms by means of adrenaline/noradrenaline eukaryotic hormones.[10]
In Gram-positive bacteria, QS regulation is usually mediated by autoinducing peptides (AIPs), that is, oligopeptides consisting of 5 – 17 amino acids, which can be linear or cyclic. The genus Staphylococcus uses the agr QS system; Streptococci use ComX, and Bacilli employ Rap system.[19] In particular, using AIPs I – IV, the agr QS system controls the expression of virulence factors (hemolysins, leukocidins, cell surface adhesins, and exoenzymes) and biofilm formation for S. aureus.[6][20]
Indole has also proved to be a signalling molecule. It affects both indole-producing and indole-non-producing bacteria. The effect depends on the type of bacterium: for example, indole increases drug resistance in Salmonella enterica and induces spore formation in Stigmatella aurantiaca.[7] Some authors also suggest that indole regulation may play a role in the adaptation of bacteria to nutrient-poor environments, in which amino acid catabolism is an important source of energy.[21] Intestinal bacteria sense the concentration gradient of indole in the intestine and thus regulate their motility, biofilm formation, and drug resistance.[10]
Some bacteria produce fatty acids as QS signalling molecules, so-called diffusible signal factors (DSFs).[22] For example, Xanthomonas campestris produces cis-11-methyldodec-2-enoic and 13-methyltetradecanoic acids; P. aeruginosa produces cis-dec-2-enoic acid; and Burkholderia cenocepacia generates cis-dodec-2-enoic acid. Diffusible signal factors perform intricate intra- and interspecies communication and also communication between different biological kingdoms. In the intraspecies communication, DSFs regulate the biofilm formation, resistance development, and production of virulence factors in Xanthomonas and P. aeruginosa. In the case of B. cenocepacia, the DSF system is connected to the AHL QS system. In the interspecies communication, DSFs of one bacterial species can affect the biofilm architecture, AI production, and production of virulence factors in a different, competing bacterial species. The suppression of C. albicans fungus, a component of the normal human microbiome, by DSFs from pathogenic bacteria, can be considered as an example of inter-kingdom communication.[23]
One more AI type in P. aeruginosa is 2-heptyl-3-hydroxy-4-quinolone (PQS). PQS-mediated QS system depends on the Las system and is a linking unit between the Las and Rhl systems.[24] This attests to a hierarchical organization of QS systems in P. aeruginosa, although under certain growth conditions and in some P. aeruginosa strains, the hierarchy is flexible.[25] It is of interest that PQS migrate not in the free soluble form, but as membrane vesicles.[24] Apart from the signalling function, PQS has antimicrobial, cytotoxic, and immunomodulatory properties[25] and also can chelate an iron ion.[26]
Special mention should be made of a QS-related phenomenon, namely, the formation of bacterial biofilms, i.e., self-regulating bacterial communities. This promotes the survival and adaptation under changing and unfavourable environmental conditions.[27-29] This process includes the following stages:
(1) adhesion of microbial planktonic cells to biotic (damaged endothelium, necrotic tissues, mucous membranes) or abiotic (catheters, prostheses, indoor walls) surface by van der Waals forces, flagella, and fimbriae;
(2) formation of the extracellular polymer matrix with the goal to form microcolonies with subsequent accumulation of multilayer clusters of microbial cells;
(3) maturation of the biofilm, in which it acquires a highly structured three-dimensional mushroom architecture pierced with channels for the transport of nutrients and autoinducers. [30] This is followed by detachment of the biofilm and dispersion of biofilm parts or single planktonic bacteria for the subsequent spreading and generation of colonies at new sites.[29]
Each of the above stages is regulated through various types of QS AIs. For example, using C4-HSL, the RhlI/R system controls the production of rhamnolipids, which promotes the twitching motility of P. aeruginosa, which is important at the adhesion stage. N-Acylated homoserine lactones also regulate the adeFGH efflux pump genes and production of lectins, necessary for the formation of the biofilm matrix. In the case of Streptococcus pneumoniae D39, by expressing the luxS gene, autoinducer AI-2 can increase the intracellular concentration of iron, which stimulates the biofilm formation, and also participates in the release of extracellular DNA, which is an important component of the matrix.[28] The agr system, which uses AIP as the autoinducer, regulates the secretion of exoproteases that induce biofilm depolymerization, which is important at the final stage of biofilm detachment. An increase in the agr gene expression increases the production of phenol-soluble modulins (PSMs), which promote biofilm maturation and dispersion.[30] Examples of various types of autoinducers are summarized in Table 1[31].
The pathogens that cause infectious diseases in humans can be controlled by affecting the communication processes between bacteria. The strategy of disrupting (or suppressing) QS systems has advantages over standard antimicrobial therapy, as it neutralizes pathogen in a non-bactericidal way. In view of the fact that most virulence factors are not essential for bacterial viability, quenching of virulence almost does not induce the development of bacterial resistance and also preserves the beneficial resident flora.[2]
Several approaches to inhibition of QS systems are distinguished
(1) inhibition by autoinducer antagonist molecules, which may be structurally similar to natural bacterial AIs, or may differ from them, while being functional mimetics. Also, both direct competitive and non-competitive antagonisms between AI and QS inhibitors are possible;[32]
(2) inactivation of AIs, which is accomplished by enzymes,[33][34] formation of host – guest complexes, e.g., using cyclodextrin,[35] or binding to monoclonal antibodies or to natural and engineered decoy receptors;[36]
(3) decrease in the AI secretion by inhibiting the appropriate synthases;[37]
(4) use of peptides to suppress the virulence of Gram-positive bacteria the QS systems of which are regulated by AIP.[31]
It should be noted that many known QS inhibitors are plant-derived compounds, since plants have co-evolved with the microbial environment and had to develop chemical mechanisms to combat bacterial infections.[38-40]
Currently, there are a few published reviews devoted to the structure – activity relationships for the secondary plant metabolites and their derivatives exhibiting antimicrobial (bactericidal and bacteriostatic) activities. The existing reviews consider almost every class of compounds: terpenes,[41][42] quinones,[43][44] coumarins,[45] stilbenes,[46] alkaloids,[47-49] curcuminoids,[50] flavonoids,[51] and phenols.[52][53] However, there are no review publications on this subject addressing particularly QS inhibitors. Meanwhile, elucidation of these dependences is highly important, because they would enable targeted search for plant-derived compounds and their analogues with enhanced efficiency, reduced toxicity, improved solubility, and acceptable pharmacokinetics that could effectively compete with antibiotics and synthetic antimicrobial drugs.
2. γ-Lactones
The γ-lactone ring is a fairly common structural moiety in natural compounds, among which there are derivatives possessing cytotoxic activity,[54] antitumour properties,[55] and antifungal activity.[56]
Currently, only one γ-lactone-based QS inhibitor has been reported in the literature. Inchagova et al.[57] isolated γ-lactones 1 and 2, analogues of AHL, from the leaves of manna gum (Eucalyptus viminalis). The γ-lactones were studied for their activity and were found to exhibit quorum sensing inhibition activity. The effective concentrations causing 50% (EC50) and 100% (EC100) inhibition of the quorum sensing-dependent biosynthesis of violacein were 2 – 4 times lower for γ-octanolactone (2) than for γ-caprolactone (1) (Table 2).
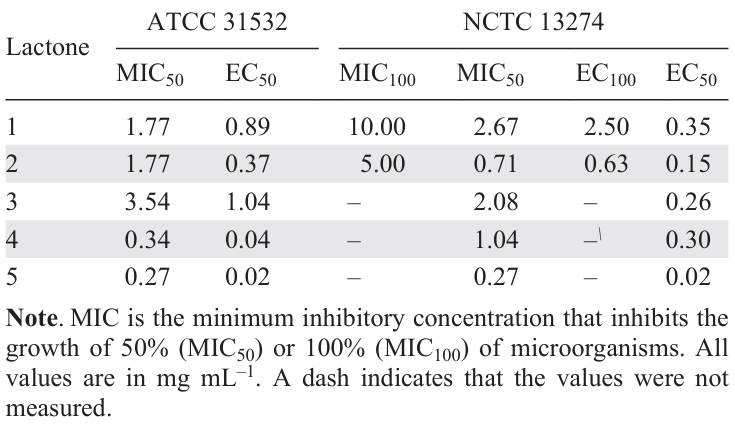
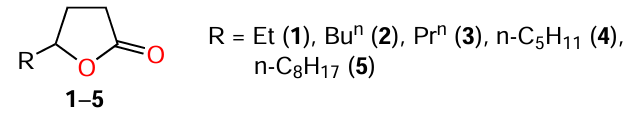
An earlier study[58] of a number of synthetic γ-lactones 3 – 5 confirmed that an increase in the length of the alkyl side chain results in higher inhibitory activity against QS (see Table 2), which is, most likely, related to increasing the structural similarity with bacterial AHLs. γ-Lactones 1 – 5 are attractive, first of all, due to their simple structure.
3. Terpenes
Despite the obvious structural similarity between γ-lactones and AHLs, most of the studied and described structural analogues of AHLs belong to terpenoids. However, due to the great structural diversity of terpenes and the complex structure of some of them (e.g., triterpenoids and sesquiterpenoids), the currently available data for establishing structure — activity relationships for compounds of this type are scarce and fragmentary.
3.1. Monoterpenes
As noted above, data on the quorum sensing inhibitory activity of terpenes are fairly scattered, which is also true for monoterpenes 6 – 13. However, certain aspects of the structure – activity relationships can be identified on the basis of published data.
The stereochemistry of natural compounds markedly influences their biological properties. For example, Ahmad et al.[59] tested the production of violacein and pyocyanin in Gram-negative anaerobic coccobacillus Chromobacterium violaceum and aerobic bacillus Pseudomonas aeruginosa and found that (+)-enantiomers of monoterpene hydrocarbons (+)-6 and (+)-8 and alcohol (+)-10 cause an increase in the violacein production. Meanwhile, (–)-enatiomers (–)-8 and (–)-10 [except for (–)-α-pinene (6)] inhibit the violacein production, which proves the presence of anti-quorum sensing activity in these compounds (Table 3).

The influence on the production of pyocyanin, a virulence factor of P. Aeruginosa,[60] follows the same pattern: the highest activity is inherent in an enantiomer of monoterpene alcohol, (–)-borneol [(–)-10].
The quorum sensing inhibition manifested by monoterpenoids depends on several structural factors.
(1) Carbon chain length.
Jaramillo-Colorado et al.[61] arrived at the conclusion that, although many monoterpene-based essential oils do not have direct structural similarity with AHLs, they still contain an analogous linear hydrocarbon substituent characteristic of short-chain (C6) derivatives and, hence, they can competitively inhibit short-chain-mediated violacein production. Meanwhile, citral (12) proved to be more effective in the inhibition of QS mediated by long-chain (C12) AHLs.
(2) Carbon chain branching
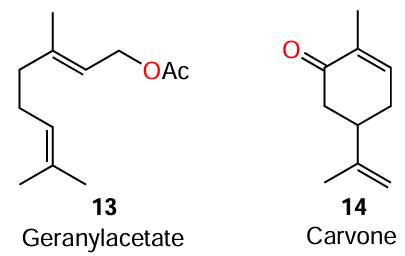
It was shown that, although geranyl acetate (13) formally has twelve carbon atoms in the molecule, it loses the similarity with long-chain AHLs because of carbon chain branching; meanwhile, it exhibits activity towards QS systems mediated by C6-AHL.[61]
(3) Size and nature of the ring in alicyclic derivatives.
Similarly to the above example, despite the presence of ten carbon atoms, limonene (8) and carvone (14) molecules are more compact owing to the six-membered ring, and this accounts for the capability of competitive inhibition of quorum sensing mediated by 3-oxo-C6-AHL.[61]
(4) Presence of a lactone moiety.
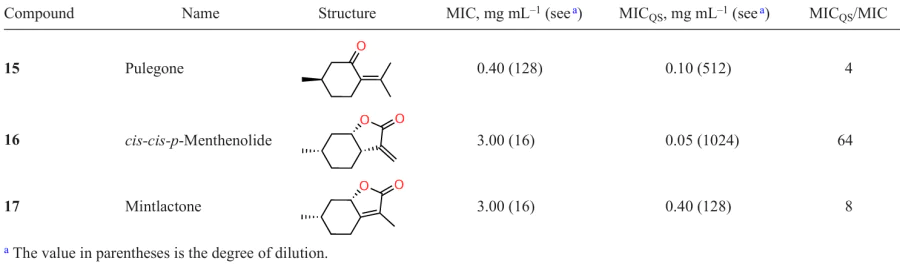
Poli et al.,[62] who investigated the biological properties of compounds 15 – 17, found out that cis,cis-p-menthenolide (16) and mintlactone 17,* which contain an unsaturated lactone ring, exhibit more pronounced anti-quorum sensing activity rather than antibacterial activity, unlike pulegone (15) containing no lactone moiety. The presence of anti-quorum sensing activity was established by assessing violacein production (Table 4). This is not surprising in view of the fact that γ-lactones described previously and monoterpenes are able to inhibit quorum sensing owing to their structural similarity to AHLs, but to different parts of AHL molecules: lactones resemble the head, monoterpenes are similar to the tail, while compounds 16 and 17 combine both AHL moieties in their molecules. Furthermore, the ratio of MICQS and MIC values is much higher for cis,cis-p-menthenolide (16) than for lactone 17 (64 and 8, respectively), indicating more efficient quorum sensing inhibition in the former case. The two compounds differ only in the position of the double bond (exocyclic or endocyclic), while exact mechanisms of the influence of this factor on the anti-quorum sensing activity are still to be elucidated.
Bound et al.[63] investigated the anti-quorum sensing activity against P. aeruginosa and C. violaceum for a number of terpene phenols (11, 18) and alcohols (19 – 21) and their glycosides, derivatives of 2,3-unsaturated 1-O-glucose (an additional letter a designates acylated glycosides and letter b implies deacetylated ones) and 2,3-dideoxy-1-O-glucose (c) (Table 5).
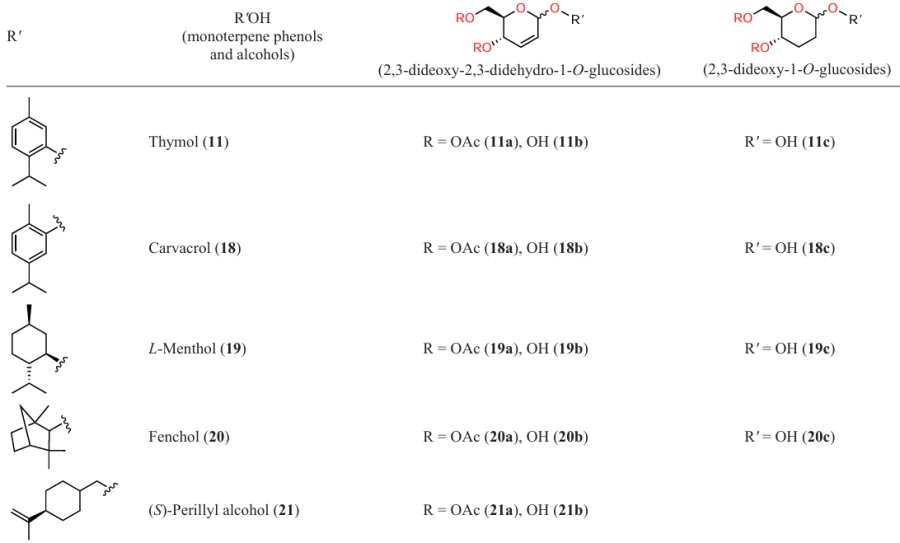
The anti-quorum sensing activity of these compounds was evaluated by testing the pyocyanin and violacein production. The authors noted two circumstances:
(1) anti-quorum sensing activity of deoxyglycosides is higher, although slightly, than that of initial terpenes almost in all cases;
(2) terpene alcohols and their derivatives (19, 20) are markedly less active than other studied compounds (Table 6).

However, the authors have not yet been able to establish the exact mechanism of QS inhibition for this type of compounds.
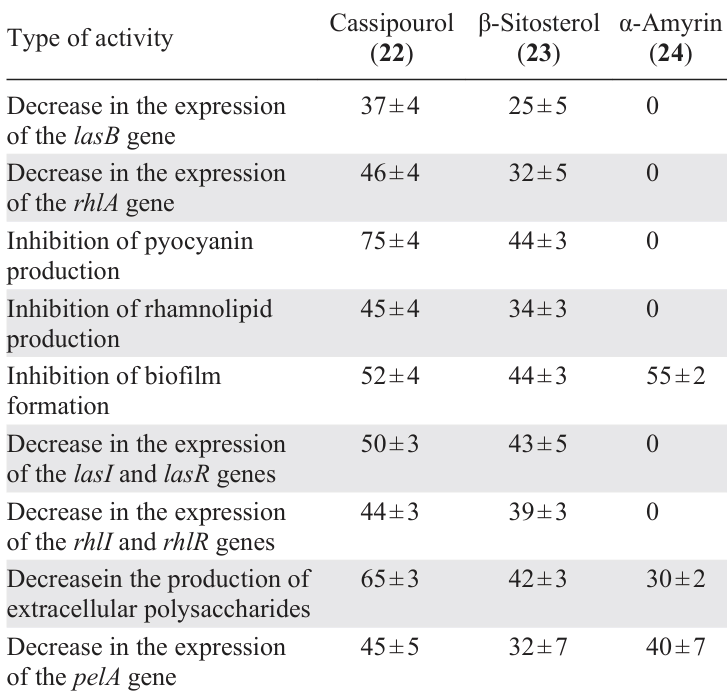
Rasamiravaka et al.[64] isolated three terpenoids, cassipourol (22), β-sitosterol (23), and α-amyrin (24), from the plant Platostoma rotundifolium. The extensive study of quorum sensing inhibition for these terpenoids included their effect on the QS-dependent expression of the lasB and rhlA genes, expression of the lasI/R and rhlI/R genes, production of virulence factors related to the quorum sensing, biofilm formation, and the ability to decrease the production of extracellular polysaccharides and suppress the expression of the regulatory pelA gene in P. aeruginosa. It is important to note that the authors showed the absence of any antibacterial activity for all three isolated compounds and found that their biological effect is attributable to quorum sensing inhibition alone, without bactericidal (bacteriostatic) action.
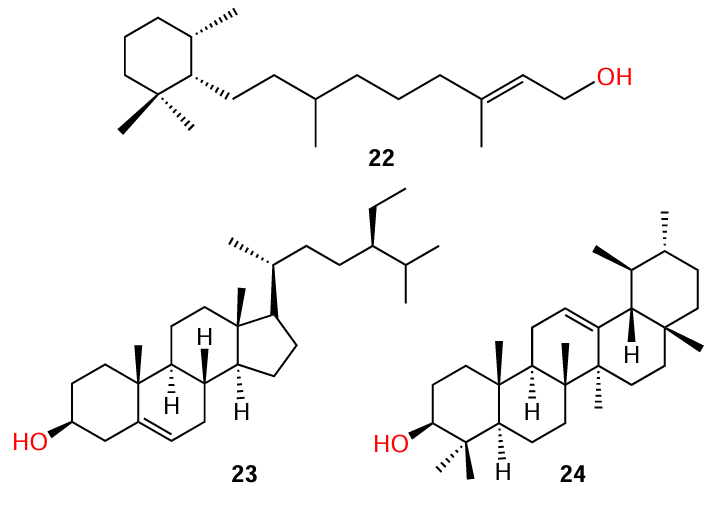
According to the authors,[64] terpenoid 22, first, showed a biological activity in all anti-quorum sensing experiments and, second, was the most effective inhibitor among the three indicated compounds almost in all cases (Table 7). The authors believe that, due to the close structural similarity to AHL, cassipourol (22) has the potential to interact with ligand-binding domains in LasR and RhlR and to compete with AHL for binding to these domains. β-Sitosterol (23) also exhibited inhibitory activity in all the experiments; however, it was less effective than compound 22. Apparently, this can be attributed to the presence of AHL-like moiety in the molecule of the former. Meanwhile, the inhibitory activity of α-amyrin (24) was observed only against the biofilm formation and against the synthesis of extracellular polysaccharides as well as in the study on the pelA gene expression.
Despite the fact that compound 24 has no structural similarity to AHL, it is a biosynthetic precursor of ursolic acid (25) (see also Section 3.2).[65] This triterpene acid is well known by its antibacterial activity[66] and the ability to inhibit the biofilm growth by a mechanism unrelated (at least, partially) to the influence on QS. This circumstance distinguishes the mechanism of action of ursolic acid from that of α-amyrin, which does not have a bactericidal activity.[67]
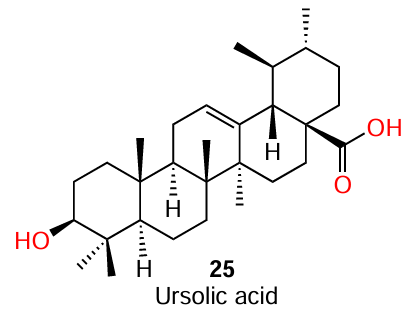
Presumably,[68] the biological activity of ursolic acid is associated with the presence of a methyl group at C(23) and a hydroxy group at C(3), which endows the molecule with hydrophilic properties and, thus, facilitates its penetration deep into the bacterial biofilm.[69] However, the particular structural moieties, which are responsible for the α-amyrin activity are still to be identified.
* This name refers to nepetalactone, one of the major components of catnip essential oil responsible for its odour.
3.2. Triterpenes
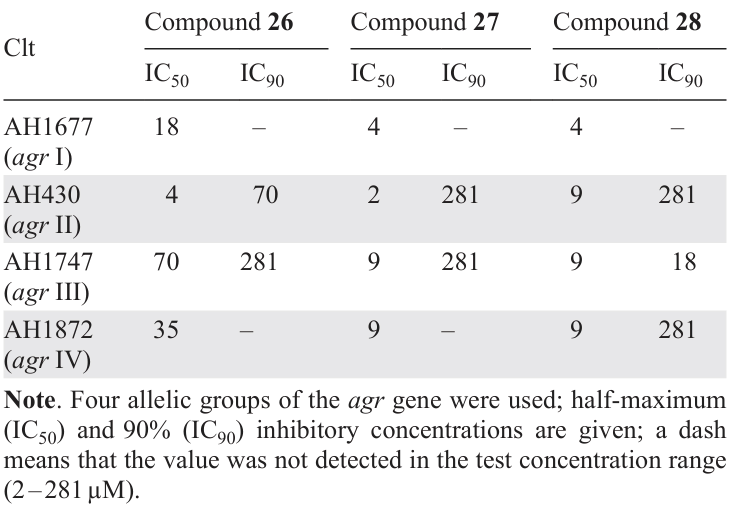
Triterpenoid acids 26 – 28, including last two having tirucallane type structure, were isolated by Tang et al.[20] from the fruit of widely used in Brazillian folk medicine, in particular to treat skin and soft tissue infections,[70] medicinal plant Schinus terebinthifolia. The authors studied for the first time the anti-QS activity of these compounds against various MRSA strains. All three compounds proved to be potent QS inhibitors: they inhibited the expression of all agr alleles (Table 8) and production of virulence factors (δ-toxin, leucocidin A, gehB lipase, nuclease), and, according to in vivo experiments, they prevented dermonecrosis. Compound 28 proved to be the most effective inhibitor in vitro, while compound 27 was most effective in in vivo assay. In the authors’ opinion, the crucial role in determining the quorum sensing inhibition belongs to the substituent at C(3), which is variable in these compounds. The presence of AHL-like moiety in compounds 27 and 28 also deserves mention.
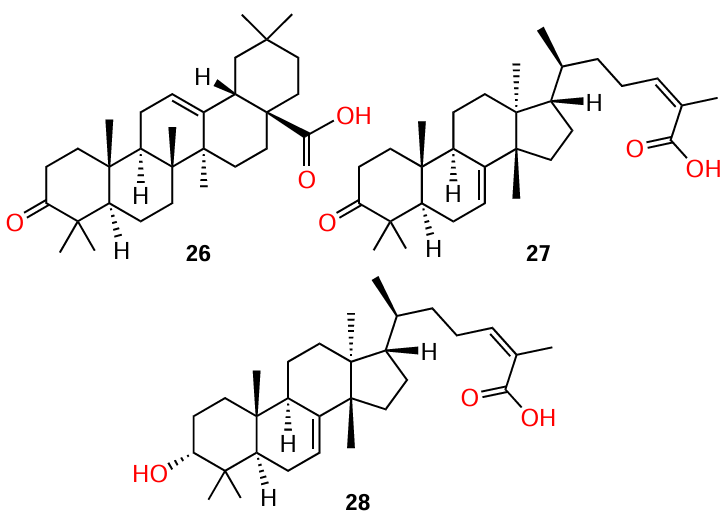
Tamfu et al.[71] isolated a group of pentacyclic triterpenoids 29 – 33 from a 70% aqueous ethanol extract of propolis of bees collecting pollen at Cameroonian grassland. Biological assays (inhibition of violacein biosynthesis, biofilm formation, and bacterial swarming) indicated that these compounds are QS inhibitors. Unfortunately, the authors were unable to elucidate the exact structure – activity relationships. However, first, the molecules of three cycloartane triterpenoid acids 29 – 31 and alcohol 32 contain an AHL-like moiety and, second, the authors hypothesized that polar substituents such as carboxyl, hydroxyl, and carbonyl groups are involved in the anti-biofilm action, as has already been mentioned above. The anti-quorum sensing properties of compound 33 are not reported in this publication.
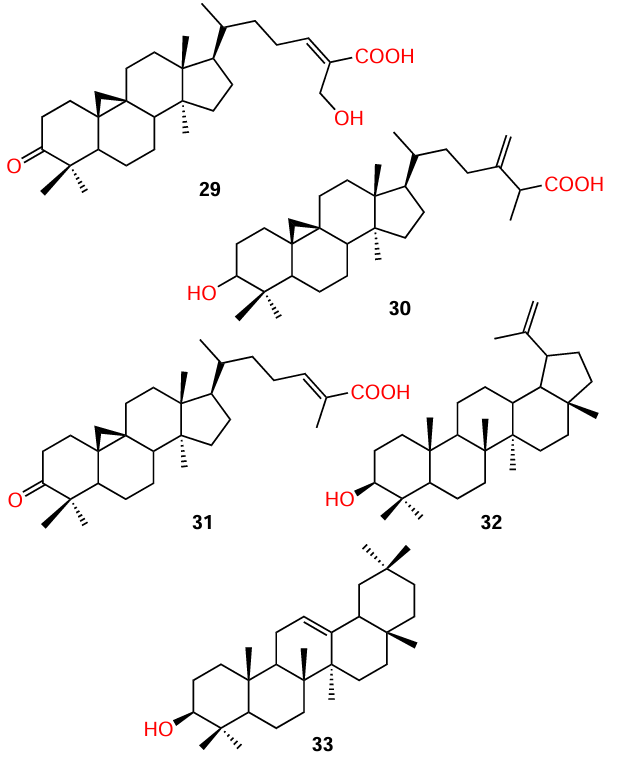
While investigating Senegalia nigrescens species, Bodede et al.,[9] in addition to isolation of known ent-kaur-15-en-18-ol (34), were able to isolate for the first time a new ent-kaurene diterpenoid, ent-kaur-15-ene-18,20-diol (35); the properties of the new product were described in detail. The isolated compounds proved to be relatively weak QS inhibitors: according to the results of the violacein inhibition assays, they demonstrated inhibition rates of 41.8% and 26%, respectively, at a concentration of 700 μg mL–1. The authors noted that the additional loading of functional groups on the kaurene pharmacophore leads to a decrease in the anti-quorum sensing activity.
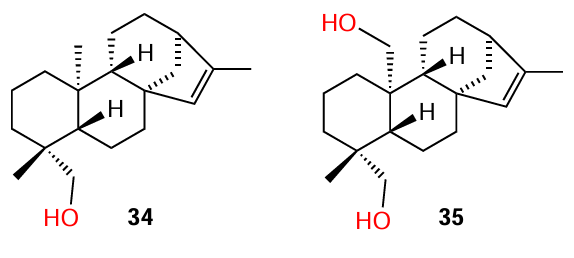
Gilabert et al.[72] isolated a number of terpenoids from the liverwort Lepidozia chordulifera, in particular, five pentacyclic triterpenoids (25, 36 – 39), three of which contained a carboxyl group, five dammarane type triterpenoids (40 – 44), and two aromadendrane sesquiterpenoids, including viridiflorol (45). The compounds were tested for biofilm formation inhibition in P. aeruginosa and S. aureus, the major nosocomial pathogens.
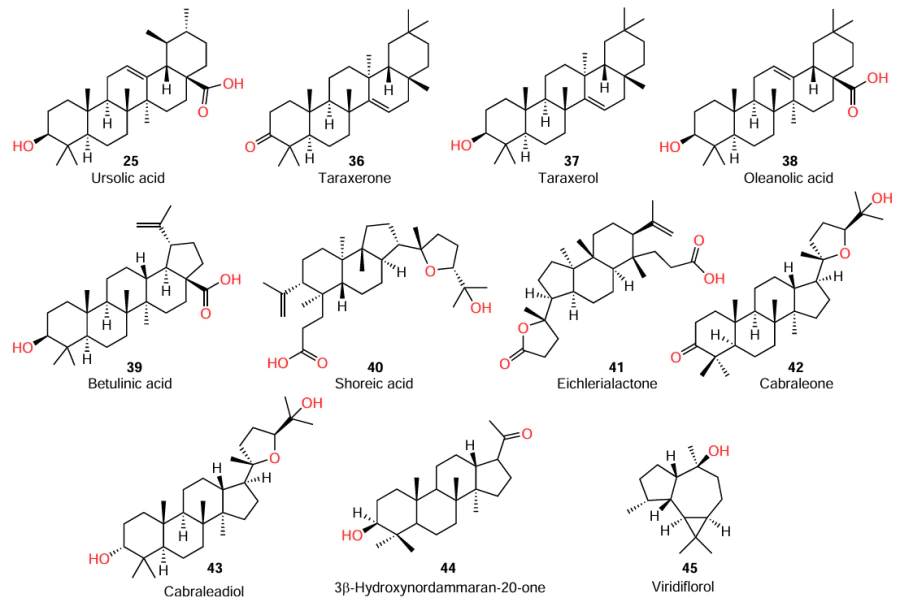
The authors identified certain aspects of the structure – activity relationships for the isolated compounds; the results are listed below (Table 9).
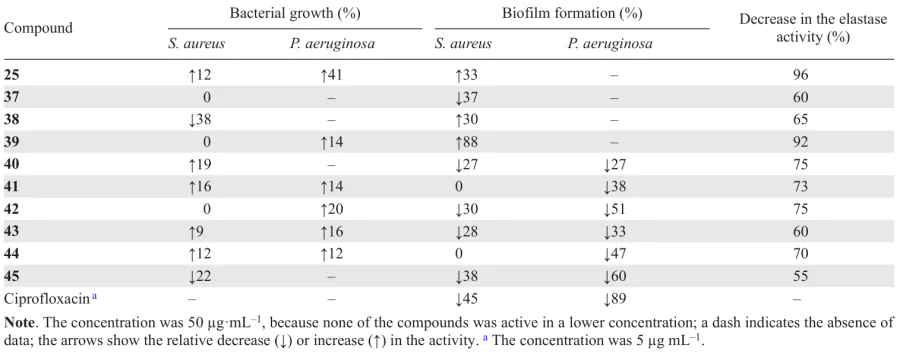
(1) Dammarane-type triterpenoids 40, 42, and 43 are able to inhibit the biofilm formation in S. aureus. A key factor for the activity is the presence of a tetrahydrofuran ring linked to a propan-2-ol group at the C(17) position of the steroid molecule. Particularly, the presence of this moiety provides the most favourable electronic and spatial conformation of the molecule.
(2) Dammarane type triterpenoids 40 – 44 also showed the most pronounced biofilm formation inhibition among compounds of this series against P. aeruginosa. Furthermore, neither the presence of a tetrahydrofuran ring at C(17) (compounds 40 – 43), nor the presence of a carbonyl or hydroxyl group at C(3) (compounds 42 – 44), nor the open steroid ring A (40, 41) had a significant effect on this type of activity. Hence, the inhibition of biofilm formation in P. aeruginosa and in S. aureus is achieved through different mechanisms.
(3) The ability to inhibit the biofilm formation in S. aureus was also found in pentacyclic triterpenoid 37 containing no carboxyl group in the molecule. Conversely, pentacyclic triterpenoids 25, 38, and 39, which contain carboxyl group, promoted the biofilm growth. This indicates that the presence of a carboxyl group is among the factors determining the activity.
(4) Apart from influencing the film growth, pentacyclic triterpenoid acids 25 and 39 were most effective inhibitors of elastase, a virulence factor of P. aeruginosa, among this series of compounds. However, it is important to note that ursolic acid (25) was markedly less active, which means that the carboxyl group at the C(17) position is not the only structural factor controlling the activity against elastase.
3.3. Sesquiterpenes
As noted above, Gilabert et al.,[72] isolated the sesquiterpenoid viridiflorol (45) from the liverwort Lepidozia chordulifera and investigated the properties of this compound. However, it was less active in the ability to control the elastase of P. aeruginosa than triterpenoids 25, 37 – 44. Nevertheless, compound 45 was active in inhibiting the biofilm formation (see Table 8 ). Previously, an anti-biofilm activity was also detected in other aromadendrane sesquiterpenoids isolated from the liverworm Porella chilensis.
Four sesquiterpene lactones (46 – 49) were isolated[13] from the plant Vernonia blumeoides of the Asteraceae family, known for a broad range of biological activity,[73] and the antibacterial properties of the lactones were described.[74] Subsequently, the authors investigated the anti-quorum sensing activity of blumeoidolides A (46) and B (47) using violacein production assay and agar diffusion assay with three biosensor systems.[13] However, QS inhibitory effect of lactones 48 and 49 could not be assessed due to their insufficient quantity.
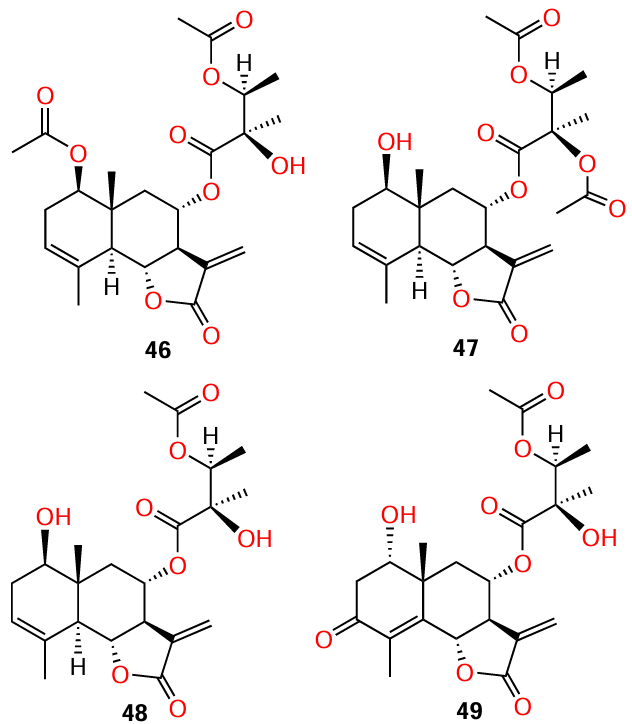
As a result of experiments, blumeoidolide B (47) proved to be a more effective QS inhibitor than blumeoidolide A (46), as it inhibited violacein production at a much lower concentration. (IC50 for compounds 46 and 47 were 1.55 and 0.055 mg mL–1, respectively). Since molecules of 46 and 47 differ only in the position of one acetyl group, the authors concluded that the presence of an acetoxy group at the side-chain C(17) atom is more favourable for the anti-quorum sensing properties than the presence of the same group at the C(1) atom of the six-membered ring. The presence of an AHL-like moiety in the molecules of both blumeoidolides was also noted.
Paz and co-workers[75][76] investigated the extracts of 15 Chilean endemic plants for the presence of QS inhibitors in them; the extracts of only three representatives (Drymis winteri J.R., Psoralea glandulosa L., and Peumus boldus) were found to be active against the reporter strain of C. violaceum ATCC 12472. Eight drimane sesquiterpenoids (50 – 57) were isolated from the bark of the most active tree, Drimys winteri.
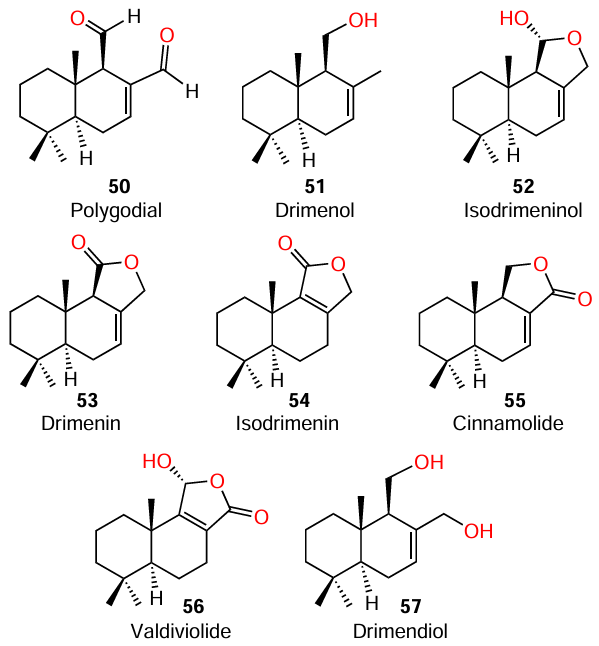
The authors were able to establish that substituents at positions 11 and 12 are determine the quorum sensing inhibition activity. Thus, polygodial (50) having aldehyde groups at positions 11 and 12 had an exceptionally pronounced bactericidal effect on the specified strain at a concentration of 100 μg mL–1. Despite the fact that five representatives of this series (52 – 56) contain, like AHLs, a lactone ring in the molecule, only compounds with a carbonyl group at position 12, cinnamolide (55) and valdivioide (56), show anti-QS activity. This is manifested as a decrease in the production of violacein pigment by 54.3% and 43.7%, respectively, at a concentration of 800 μg mL–1. Lactones with a carbonyl group at position 11, drimenin (53) and isodrimenin (54), proved to be inactive. According to the results of the authors, drimendiol (57), prepared by means of the reduction of polygodial (50) and containing hydroxyl groups at positions 11 and 12, had a more pronounced QS inhibitory activity than a,b-unsaturated lactones, providing 70% inhibition of violacein production at a concentration of 800 μg mL–1.
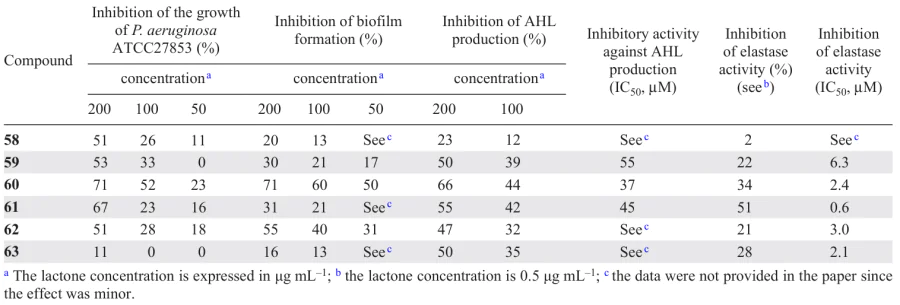
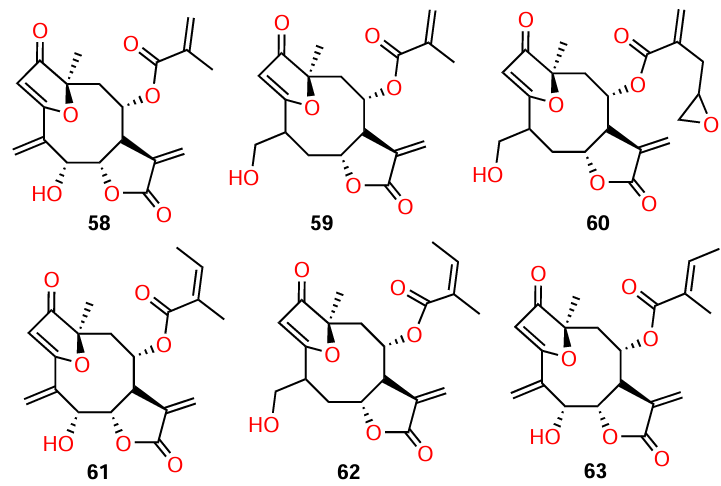
Amaya et al.[77] isolated the sesquiterpene lactones 58 – 63 from the plant Centratherum punctatum. Among these compounds, goyazensolide type lactones (59, 60, and 62) showed a higher quorum quenching activity: inhibition of biofilm formation, AHL production, and P. aeruginosa elastase activity compare to isogoyazensolide type lactones (58, 61, and 63) (Table 10). The activity of the former proved to be comparable with that of other sesquiterpene lactones of higher plants[78] and synthetic N-sulfonyl homoserine lactones described by Castang et al.[79]
4. Phenols
Apart from thymol (11), which was mentioned above, structural analogues of AHLs were found among phenol derivatives; an example is zingerone (64) isolated from ginger (Zingiber officinale). This plant is an extremely popular spice and has antimicrobial properties,[80][81] which makes it applicable in folk medicine.[82][83] Kumar and co-workers[84][85] suggested that there may be QS inhibitors among biologically active ginger components 64 – 68. Indeed, zingerone (64), a major aromatic component isolated from the dried ginger root, showed a high level of quorum sensing inhibition in P. aeruginosa PAO1 strain: in the presence of this phenol, the numbers of so-called swimming, swarming, and twitching phenotypes were found to decrease. In addition, the authors observed decreasing biofilm formation and production of virulence factors such as rhamnolipid, elastase, protease, pyocyanin, and cell-free and cell-bound hemolysin.[85]
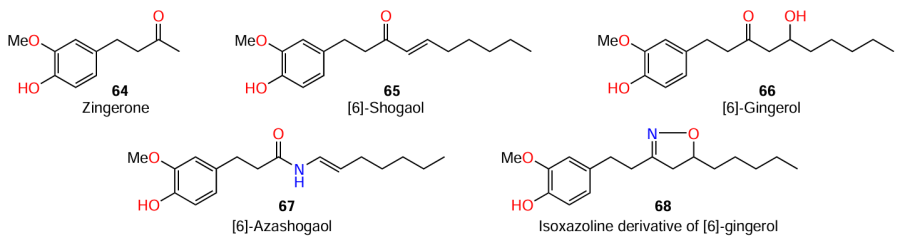
To establish the structure – activity relationships, the authors also investigated some derivatives of zingerone.[84] According to qualitative (agar well diffusion method) and quantitative (MIC determination by broth microdilution test, and pyocyanin and violacein pigment assay) methods, zingerone (64) proved to be the least active QS inhibitor of the bacteria C. violaceum and P. aeruginosa, while [6]-azashogaol (67) showed the highest activity. In the latter case, the zone of inhibition was 17 mm (Table 11) and decrease in the pyocyanin production was 90%.

Considering these results, the authors elucidated the following regularities:
(1) As the length of the alkyl side chain increases, the structural similarity of the compounds to the AHL tail increases and, hence, their anti-quorum sensing activity is enhanced. That is why zingerone (64), which has the shortest alkyl substituent in this series, has the lowest activity, while the most effective inhibitor, [6]-azashoagol (67), contains the longest alkyl side chain;
(2) the anti-quorum sensing activity is additionally enhanced upon the introduction of an amide bond between the alkyl side chain and the phenol moiety, whereas the heterocyclic linker (isoxazoline, in the case of compound 68) does not have a significant effect on QS inhibition.
5. Tannins
The QS inhibitors described in the previous Sections were either virtually complete analogues of AHLs or contained AHL-like moieties in the molecules. However, most of compounds that exhibit QS inhibitory activity do not have structural similarity to AHLs, but mimic their action, i.e., they act as AHL mimetics. These QS inhibitors may be either simple small molecules such as tannins [in particular, methyl gallate (69)] or more complex structures, including fused ones.
Tannins, which belong to the class of polyphenols, have tanning properties due to their ability to bind to proteins, polysaccharides, and biopolymers. For this reason, these compounds are discussed in this separate Section.
Hossain et al.[86] investigated series of tannins, among which only methyl gallate (69) exhibited quorum sensing inhibitory activity against C. violaceum and P. aeruginosa. Since the unsubstituted precursor, gallic acid (70), was inactive,[87][88] the authors assumed that the presence of the gallic moiety in the molecule was insufficient for the compound to exhibit anti-quorum sensing activity.
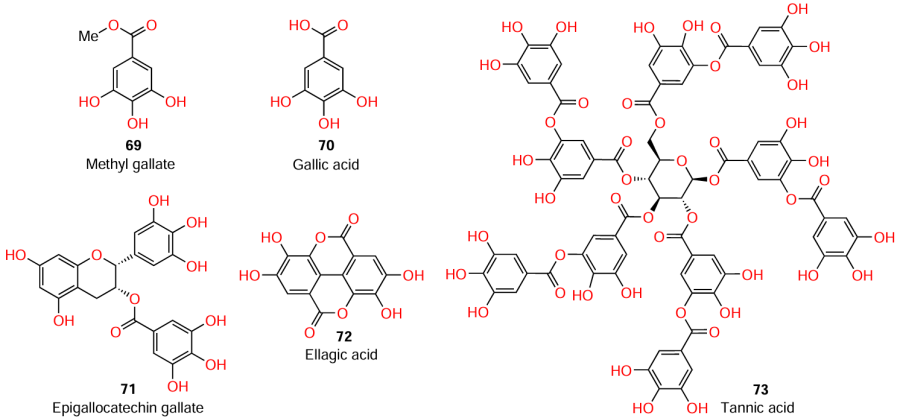
This hypothesis was further developed by Pimenta et al.[89] Since it is the methyl ester group that endows methyl gallate with the activity, it can be assumed that replacement of the hydrogen atom in the gallic acid carboxyl group by other moieties would afford new derivatives possessing quorum sensing inhibitory activity. This hypothesis is indirectly confirmed by the fact that structural analogues of gallic acid such as epigallocatechin gallate (71), ellagic acid (72), and tannic acid (73) act as QS inhibitors in case of P. aeruginosa and some other pathogens.[90-97]
6. Curcuminoids
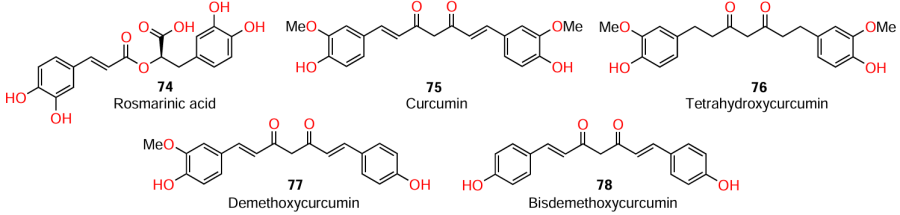
Curcuminoids, secondary metabolites of the turmeric plant (Curcuma longa L.) are widely used as food colorants due to their bright yellow colour. A known AHL mimic is rosmarinic acid (74), a functional analogues of C4-AHL.[98][99] Some authors noted[6][98] that structural analogues of rosmarinic acid, i.e., linear compounds with aromatic groups at the ends of the molecule, also mimic AHL and, hence, inhibit QS.
The source of curcumin (75), common turmeric (Curcuma longa L.), is a popular Asian spice with a pronounced antimicrobial activity.[100] As has already been noted above, compound 75 exhibits quorum sensing inhibitory activity against S. aureus, but has no antibacterial activity. In particular, Park et al.[101] found that curcumin and its derivatives inhibit one of the most important virulence factors of Gram-positive bacteria, the sortase A enzyme (SrtA). As regards the structure – activity relationships, the authors noted that for the presence of inhibitory activity, the linker should contain double bonds and methoxy groups in the aromatic rings as pharmacophore elements. The former ensures the structural rigidity, because, otherwise, the molecule loses the linearity and, hence, the activity, as can be seen in relation to tetrahydroxycurcumin (76). For this reason, curcumin (75) is the most potent inhibitor of SrtA among curcuminoids, characterized by IC50 = 13.8 ± 0.7 mg mL–1, while the IC50 values for demethoxycurcumin (77) and bisdemethoxycurcumin (78) are 31.9 ± 1.2 and 23.8 ± 0.6 mg mL–1, respectively.[101]
7. Cinnamaldehyde and its analogues
Considering the data from the previous Section, one can state that the presence of an α,β-unsaturated carbonyl moiety in the molecule of a natural compound or its synthetic analogue may be the crucial factor to determine the quorum sensing inhibition. In this context, it is worthy to note a study by Brackman et al.,[102][103] who addressed QS inhibition by cinnamaldehyde (79) and a broad range of its derivatives (80 – 119) against vibrions, that are, Vibrio pathogens (V. harveyi, V. anguillarum, and V. cholerae).* According to the results of the authors, no anti-quorum sensing activity was inherent in 3-phenylpropionaldehyde (111), 3-phenylpropionic acid (112), cinnamic alcohol (115), methyl cinnamate (116), pent-2-enoic acid (97), or 5-methylhept-2-en-4-one (100) (Table 12). The activity of a number of derivatives exceeded that of cinnamaldehyde; examples are some halogenated compounds [3,4-dichlorocinnamaldehyde (87), 2,3,4,5,6-pentafluorocinnamaldehyde (94) and 4-chloro-3-trifluoromethylcinnamaldehyde (92)] and aldehydes: (E)-non-2-enal (94), (E)-dec-3-en-2-one (98), (E)-non-3-en-2-one (99), (E)-oct-3-ene-2-one (104), (E)-oct-2-enal (105), (E)-dec-2-enal (107), (E)-undec-2-enal (108), and (E)-dodec-2-enal (109).
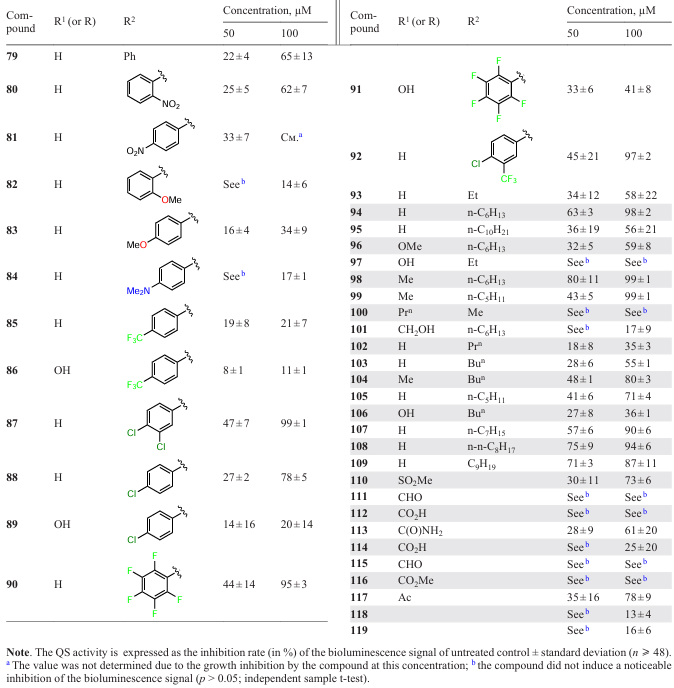

Based on the results of primary screening, compounds were selected for studying their ability to inhibit the protease activity and reduce the biomass of biofilms. It was found that virtually all compounds of this series were active in all the experiments for all three types of microorganisms (Table 13).
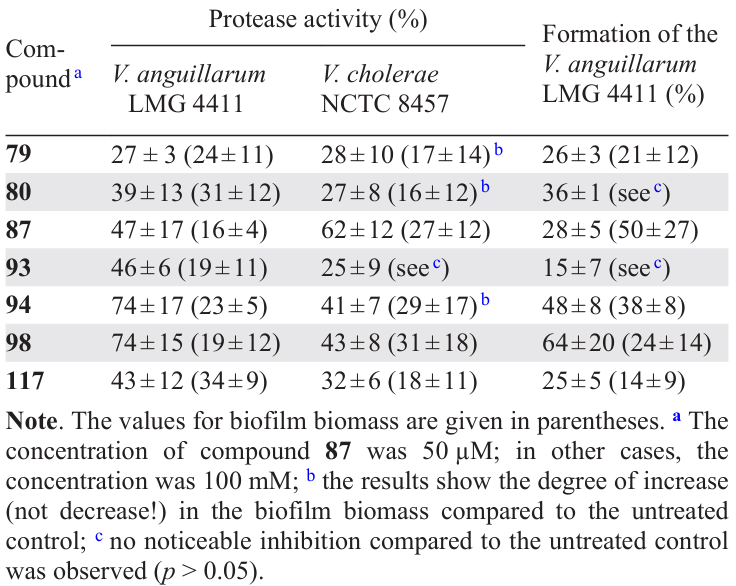
The authors suggested that the mechanism of quorum sensing inhibition by cinnamaldehyde (79) and its analogues involves reaction with the LuxR protein, which results in a change in the conformation of the last one and, thus, its lower ability to bind to DNA. In this case, QS inhibitors bind to LuxR through the Michael addition reaction. In view of the obtained data, the authors came to the following conclusions regarding the structure – activity relationships:
(1) compounds that are able to participate in the Michael addition reaction are active, while replacement of the aldehyde group by a carboxyl group reduces the ability to inhibit QS;
(2) compounds containing no carbon – carbon double bond are inactive;
(3) the inhibitory effect depends strongly on the nature and degree of substitution of the aromatic ring (e.g., the presence of electron-withdrawing substituents increases the activity);
(4) substituents providing a larger π+σ sum, where π is the Hansch constant and σ is the Hammett constants, lead to a higher inhibitory effect.
* This Section presents also aliphatic analogues of cinnamaldehyde described by the same authors.
8. Stilbenes
As mentioned above, the presence of an ethylene bridge in the molecules containing two aromatic moieties is an important factor for the anti-quorum sensing activity. The transition from biphenyl (120) and diphenylmethane (121) to derivatives with an ethylene linker leads to a large class of stilbenes. Lee et al.[104] carried out a detailed study of cis-stilbene (122), trans-stilbene (123), oxyresveratrol (124), trans-resveratrol (125), and quite a few structurally similar compounds. The compounds were evaluated as QS inhibitors against S. aureus by measuring the antivirulence activity, that is, anti-hemolytic and anti-biofilm activities. It is noteworthy that among the test compounds, only trans-stilbene exhibited both types of antivirulence activity against the model strain S. aureus ATCC 6538 with a dose-dependent effect. Compounds 123 and 125 at concentrations of 10 mg mL–1 prevented hemolysis by more than 85 and 90%, respectively, and showed anti-biofilm effect against MRSA. Meanwhile, trans-resveratrol 125 had only anti-hemolytic activity against the indicated strain, but possessed both types of activity against MRSA with a dose-dependent effect.
According to Lee et al.,[104] the anti-biofilm activity of compounds is not associated with the inhibition of bacterial growth. As a result, the following conclusions can be drawn regarding the structure – activity relationships:
(1) the anti-hemolytic and anti-biofilm effects require the presence of a stilbene moiety with a trans-C=C bond. Compounds that do not contain a C=C bond or have a cis-C=C bond either show no antivirulence activity [e.g., biphenyl (120) and diphenylmethane (121)] or exhibit oly one type of activity;
(2) the number and arrangement of hydroxyl groups in the benzene ring affect both the presence of anti-hemolytic activity [e.g., compound 124, although complies with the structural requirements indicated in (1), still did not significantly influence the hemolysis] and the selectivity to various pathogen strains (in this particular case, Staphyloccocus strains).
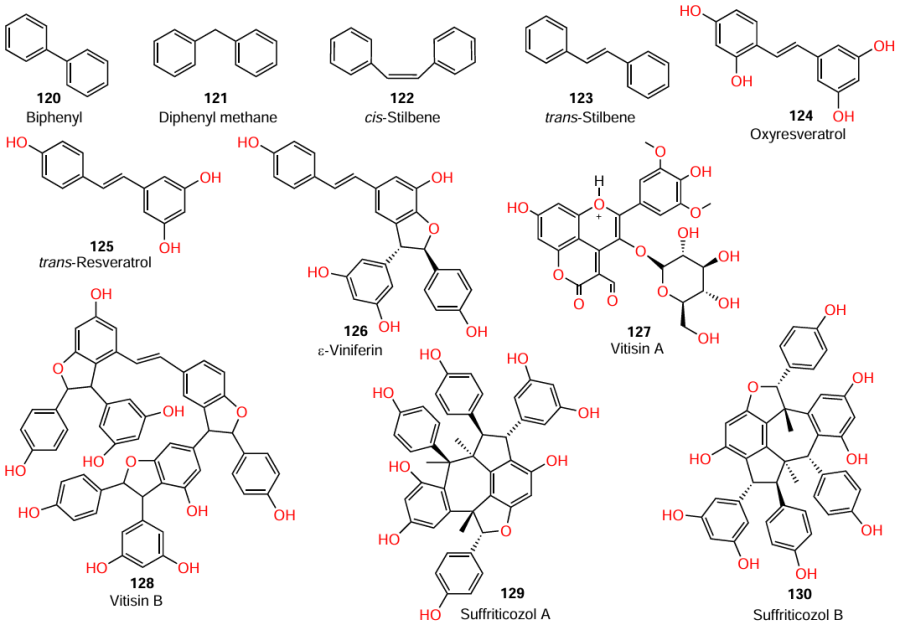
It is also important that resveratrol oligomers, ε-viniferin (126), vitisin A (127), vitisin B (128), suffriticosol A (129), and suffriticosol B (130), also have anti-hemolytic activity against S. aureus, with tetramer 128 being the most active out of the five compounds, providing > 70% and > 95% inhibition of hemolysis at concentrations of 1 and 2 μg mL–1, respectively.
Cho et al.[105] showed that, apart from hemolytic activity against S. aureus, ε-viniferin (126) also has the anti-biofilm activity against Gram-negative microorganisms: at a concentration of 50 mg mL–1, this compoundi inhibited the biofilm formation of P. aeruginosa by 82%, while the biofilm formation by E. coli was inhibited by 98% at 10 mg mL–1.
Another resveratrol tetramer that exhibited anti-quorum sensing activity [inhibition of type III secretion systems (T3SS)] against Gram-negative microorganisms, P. aeruginosa and Yersinia pseudotuberculosis, is (−)-hopeaphenol (131).[106] At concentrations from 3.3 to 100 mM, compound 131 exhibited dose-dependent and irreversible inhibition of the secretion of Yops (Yersinia outer proteins), which are effector proteins of Yersinia pseudotuberculosis. At the same concentration, this tetramer also inhibited T3SS of the P. aeruginosa superbug.
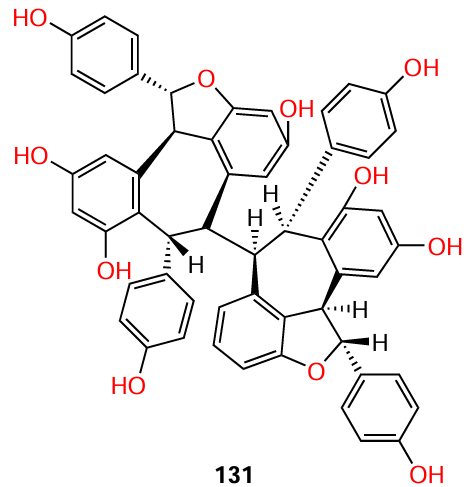
It is worthy to note that other stilbenes are also markedly inhibited QS in Gram-negative bacteria. In particular, while establishing the structure – activity relationship in the series of stilbenoids, Sheng et al.[107] chose the P. aeruginosa PAO1 strain and C. violaceum CV026 indicator strain as the pathogens. According to violacein production assay, only stilbenes 124, 125, and 132 were found to be active: at a concentration of 100 mM, they substantially decreased the swarming motility, while at 400 mM, they decreased the pyocyanin production by 59.3, 37.1, and 50.2%, respectively. In addition, these compounds reduced the expression of the lasI, lasR, rhlI, and rhlR genes (in particular, the lasI gene expression decreased two-, five-, and three-fold, respectively).
Analyis of the pattern of presence or absence of anti-quorum sensing in a series of ten stilbenes and their analogues (124, 125, 132 – 139) provides the conclusion that structural requirements for stilbenes that inhibit QS in Gram-negative microorganisms are similar to those for Gram-positive microorganisms, namely:
(1) the presence of a double bond is necessary (1,2-dierylethanes 134 – 137 and phenanthrenes 138 and 139 do not possess an anti-QS activity);
(2) the presence of hydroxyl groups at positions 3' and/or 5' is also a significant factor: the replacement of hydroxyl groups by methoxy groups results in a loss of activity, while the introduction of a hydroxyl group in position 2' decreases the inhibition rate.

The above assumption about the influence of the number and location of hydroxyl groups on the selectivity to various Gram-positive microorganisms is supported by the fact that among structurally similar compounds 122, 123, 125, 132, and 140 – 148, no anti-biofilm activity against Streptococcus mutans was observed for unsubstituted stilbenes 122 and 123, amino derivative 140, or glycoside 148, in contrast to active trans-resveratrol (125) and some other substituted stilbenes (Table 14).[108]
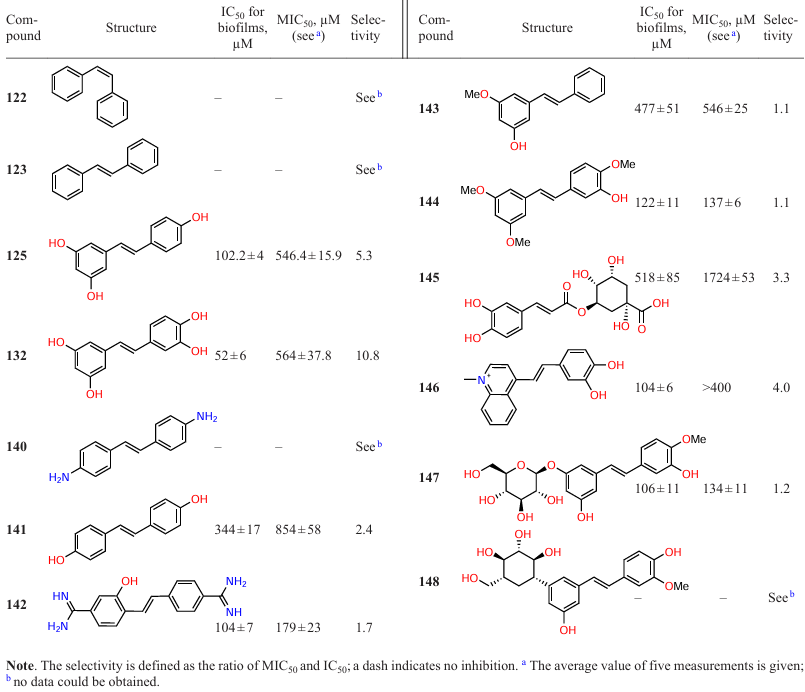
A study by Nijampatnam et al.[108]sheds light on some aspects of the structure – activity relationship for stilbenes. In particular, regarding the importance of hydroxyl groups in the molecule, the authors found that the dihydroxy analogue 141 of amino derivative 140 with similar regiochemistry of substituents has anti-biofilm activity (IC50 = 344 μM), while diamidine 142, which also contains an ortho-hydroxyl group in one benzene ring, inhibits the biofilm growth with IC50 = 104 μM.
As regards the effect of presence and regiochemistry of the hydroxyl substituents in trans-resveratrol and its derivatives, it was found that the absence of hydroxyl groups in one of the benzene rings leads to a decrease in the anti-biofilm activity and selectivity. Indeed, IC50 for 3-hydroxy-5-methoxystilbene (143) is 477 mM, whereas piceatannol (132), which differs from trans-resveratrol (125) only by the presence of an additional hydroxyl group, shows, conversely, a greater anti-biofilm activity (IC50 = 102 mM vs. IC50 = 52 mM, respectively) and much higher selectivity (10.8) to S. mutans. At the same time, compound 132 does not inhibit the growth of S. sanguinis and S. gordonii films, which also attests to exceptionally high selectivity of action. However, this effect on the selectivity cannot be attributed to particularly the hydroxyl group in the meta-position relative to the ethylene moiety, since trimethoxy derivative 144 is only slightly inferior in the anti-biofilm activity (IC50 = 122 μM), but has a significantly lower selectivity. A similar trend was observed for the methylated glycoside piceatannol analogue 147 (IC50 = 106 μM). It is of interest that compound 148, which is structurally similar to glycoside 147, but does not contain a C–O bond with the carbohydrate moiety, completely loses the anti-biofilm properties. All of the above factors indicate that the presence of four hydroxy groups in definite positions of piceatannol benzene rings is important for high antibacterial activity and selectivity.
It is noteworthy that some structure modifications are still admissible. Thus, when two hydroxyls are present in only one benzene ring and one aromatic moiety is replaced by a cycloalkyloxycarbonyl moiety or an N-methylquinoline ion, compounds 145 and 146 show fairly good biofilm growth inhibition activity (IC50 = 518 μM and IC50 = 104 μM, respectively), but there are still no published data on the structure – activity relationships for these compounds. An important remark is that the anti-biofilm activity of piceatannol is associated with the inhibition of Gtfs, a key virulence factor of S. mutans, rather than with the bactericidal action.
Piotto et al.[109] synthesized a number of azobenzenes as stilbene analogues 149 – 153. All compounds behaved as QS inhibitors, namely, they showed an anti-biofilm activity (without bactericidal action) against S. aureus.

Compound 152 proved to be somewhat less effective: it inhibited the biofilm formation when used at a concentration of 30 μg mL–1, while the other compounds of this series showed the same effect at concentrations of only 25 μg mL–1. Nevertheless, it can be concluded that replacement of the carbon atoms by nitrogens at the stilbene double bond does not induce the loss of antibacterial activity against S. aureus. Moreover, these compounds are fairly easy to obtain and, according to the preliminary calculations, they are expected to have low hepatotoxicity.
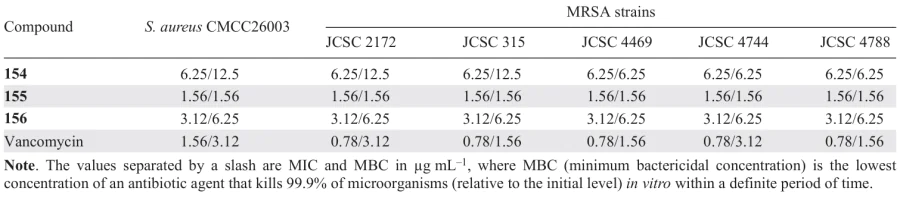
Apart from trans-stilbene and resveratrol, other natural stilbenes were also found to exhibit anti-hemolytic and anti-biofilm activity against various S. aureus. Particularly, cajaninstilbene acid (154), longistylin A (155), and longistylin C (156) isolated by Li et al.[110] from the leaves of pigeon pea (Cajanus cajan). The attention of the authors was attracted by the plant itself, as it was repeatedly mentioned in the literature that longistylin A and its derivatives are the main components of aqueous ethanol extracts of pigeon pea leaves[111][112] and anti-MRSA activity of this compound has been reported.[113]
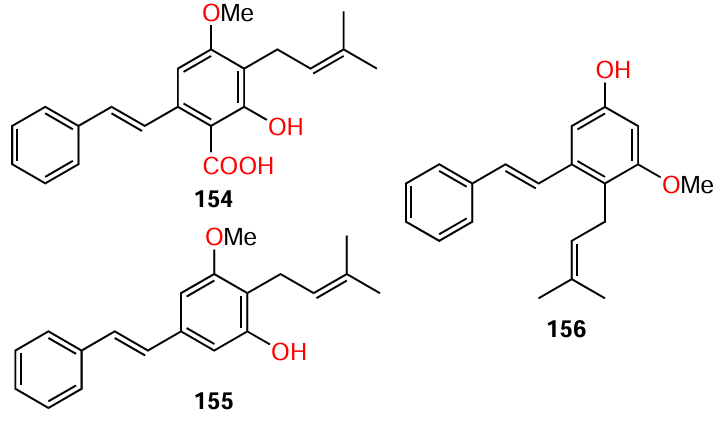
When used at a concentration equal to 1/4 of MIC (Table 15), all the stilbenes 154 – 156 markedly inhibited the adhesive properties of the test strain and MRSA biofilm formation and suppressed, to various extents, the expression of adhesion genes (icaA, icaD, clfA, and clfB) and virulence genes (AgrA, RNAIII, hla) and the prodution of α-hemolysin (a virulence factor). Among the three stilbenes, the highest anti-hemolytic activity and the most pronounced suppression of adhesion and virulence gene expression were found for cajaninstilbene acid (154).
Hunag et al.[114] synthesized 24 cajaninstilbene acid derivatives 157 – 161 and studied the structure – activity relationships in relation to the anti-biofilm activity for P. aeruginosa bacterium. Of this series, only compounds 158g,m,o considerably inhibited the biofilm formation (Table 16), with 158o being most active: the inhibition rate was 50% at a concentration of 50 mM. The other compounds either had a weak inhibitory effect or, conversely, promoted biofilm formation. Compound 158o also inhibited the expression of las and pqs systems, which, in turn, resulted in a decrease in the production of virulence factors: elastase, pyocyanin, and pyoverdine.
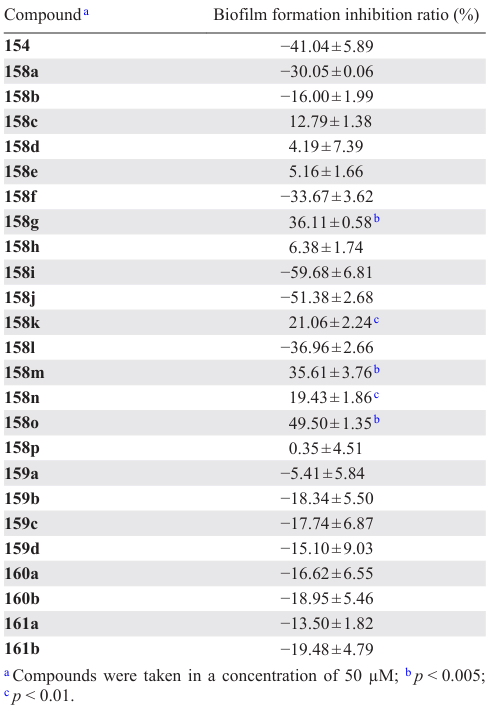
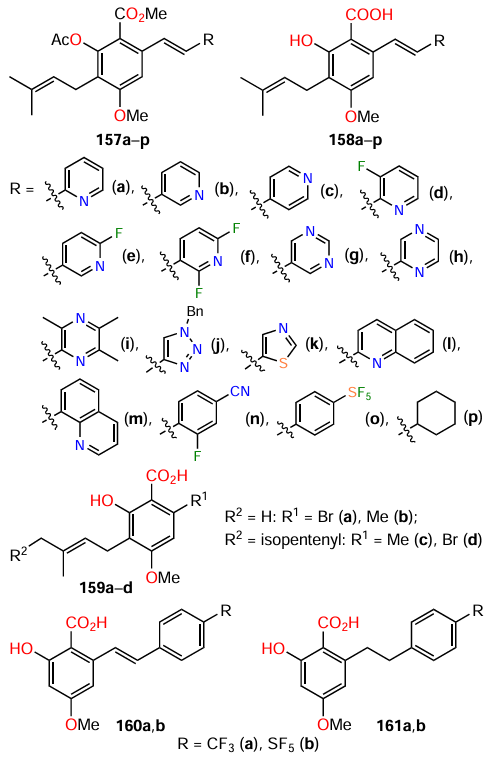
The authors drew the following conclusions:
(1) the integrity of the structure of cajaninstilbene acid (structures 158a – p) is a necessary condition for the activity to be manifested; simplified analogues, for example, those containing geranyl, isopentenyl, trifluorophenyl, or phenylsulfur pentafluoride as side substituents, did not exhibit inhibitory activity;
(2) the introduction of a pyridine or pyrazine ring into the molecule afforded compounds with a very weak or no anti-QS activity, while the presence of a pyrimidine ring markedly increased the activity of these derivatives;
(3) the quinoline ring itself is not a crucial factor determining the activity; the substituents in the ring play a much greater role, but the authors have not yet been able to identify the exact relationship.
9. Coumarins
Coumarin (2H-1-benzopyran-2-one) (162) is formally the o-hydroxycinnamic acid lactone; therefore, it is not surprising that coumarin has anti-quorum sensing properties.[115] It is known that the benzopyran moiety is present in the las and pqs systems of P. aeruginosa QS inhibitors.
Lee et al.[116] investigated the quorum sensing inhibition by a number of natural coumarins 162 – 169 against the E. coli O157:H7 strain. The compounds that proved to be most active include coladonin (163), coumarin (162), and umbelliferone (169): 80, > 80, and 90% inhibition of biofilm formation, respectively, was observed in the presence of these compounds at 50 mg mL–1 concentration; furthermore, the compounds showed no bactericidal activity. In addition, coumarins 162, 166, and 169 reduced the expression of csg operon genes (csgA and csgB) and motility genes (flhD and motB), while esculetin (166) additionally suppressed the expression of the stx2 gene of Shiga-like toxin II.
The authors identified the following patterns for the dependence of anti-quorum sensing on the structure of coumarins:
(1) the introduction of a hydroxyl group in position 4 or 8 or introduction of two hydroxyl groups in positions 6 and 7 decreases the activity, while hydroxylation to position 7 has the opposite effect;
(2) replacement of the hydroxyl group at position 6 by the methoxy group or introduction of the sesquiterpene moiety in position 7 does not influence the inhibitory effect;
(3) the introduction of a carboxyl group in position C(3) decreases the activity.
Data on the structure – activity relationship partially similar to the results of Lee et al.[116] were obtained by Deryabin et al.[117] According to the violacein production assay, among coumarins 162, 170, and 171, 7,8-dihydroxy-4-methylcoumarin (171) was most active (Table 17). Relying on this result, the authors concluded that hydroxylation of position 7 actually increases the inhibitory activity. However, the difference between the results of these two studies is that in the latter case, the introduction of two hydroxyl groups did not decrease the activity compared to that of unsubstituted coumarin, but, conversely, had the opposite effect.
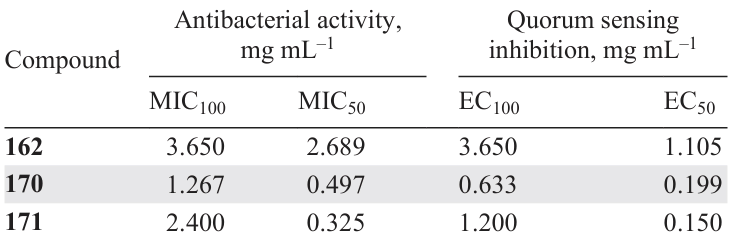
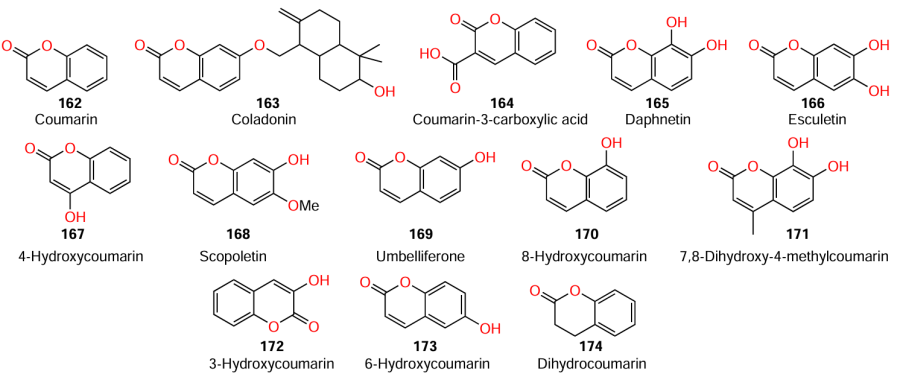
D’Almeida et al.[118] used coumarins 162, 166, 167, 169, and 172 – 174 to carry out a study similar to the above one, but using C. violaceum ATCC 12472 and P. aeruginosa ATCC 27853 strains. According to the results of both the violacein production bioassay and experiments on the inhibition of P. aeruginosa biofilm formation, 4-hydroxycoumarin (167) and dihydrocoumarin (174) were the least active. Meanwhile, seven compounds, 4-hydroxycoumarin (167), dihydrocoumarin (174), 6,7-dihydroxycoumarin (171), 7-hydroxycoumarin (169), coumarin (162), 6-hydroxycoumarin (173), and 3-hydroxycoumarin (172) at a concentration of 100 mg mL–1, decreased the biofilm formation by 20, 18, 63, 61, 53, 46, and 38%, respectively. The lowest inhibitory effect against elastase was inherent in dihydrocoumarin (174), while the inhibition rates for 7-hydroxycoumarin (169), coumarin (162), 3-hydroxycoumarin (172), 6-hydroxycoumarin (173), 4-hydroxycoumarin (167), 6,7-dihydroxycoumarin (166), and dihydrocoumarin (174) at a concentration of 20 mg mL–1 were 52, 47, 45, 43, 37, 33, and 3%, respectively. Interestingly, as opposed to the results reported by Lee et al.,[116] bis(hydroxy) derivative 166 proved to be most active in this series. The authors also concluded that hydroxyl groups in the aromatic ring and a double bond in the pyrone ring are the necessary conditions for the presence of anti-biofilm activity. The presence of a hydroxyl group at C(3) is required for elastase inhibition.
These disparate conclusions drawn by various research groups suggest that in the case of coumarins, the specificity, in particular species specificity and quorum sensing inhibition are determined by the position and number of hydroxyl groups.
10. Quinones and hydroquinones
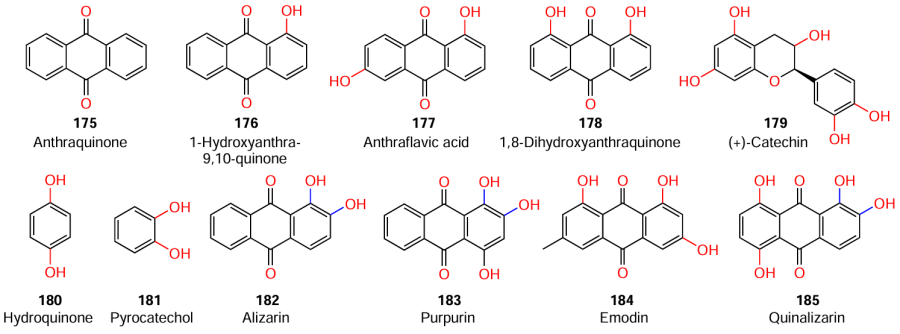
The published data on the structure – activity relationships for quinones are mainly concerned with their antibacterial activity.[43][44] The only mention of the action of quinones as QS inhibitors was made by Lee et al.,[119] who investigated the potential of these compounds against S. aureus. Among quinones and their derivatives (175 – 185), alizarin (182), emodin (184), purpurin (183), and quininalizarin (185) at a concentration of 10 mg mL–1, inhibited the biofilm formation and decreased the hemolytic activity of S. aureus MSSA 6538 by ≥70% in comparison with control samples. According to the conclusions made by the authors, important factors for the anti-biofilm activity are the presence of both the anthraquinone core and two hydroxyl groups at C(1) and C(2) atoms. The introduction of additional hydroxyl groups has only a minor effect on the inhibitory activity of quinones.
11. Flavonoids
Flavones (2-phenyl-4-chromenones, α-phenylchromones) are α,β-unsaturated heterocyclic ketones derived from γ-pyrone that are widely encountered in plant sources, flowers, and brightly coloured fruits and vegetables.[120] It is not surprising that QS inhibitors have been found among compounds of this class.
Paczkowski et al.[121] used phloretin (186), chrysin (187), and naringenin (188) to demonstrate the action of flavones as non-competitive inhibitors of LasR and RhlR. The authors found that each of the three flavonoids has two hydroxyl substituents in positions 5 and 7 of ring A. However, unlike compounds 187 and 188, the first-mentioned compound 186 does not have a rigid structure provided by a chromanone or chromenone ring C.
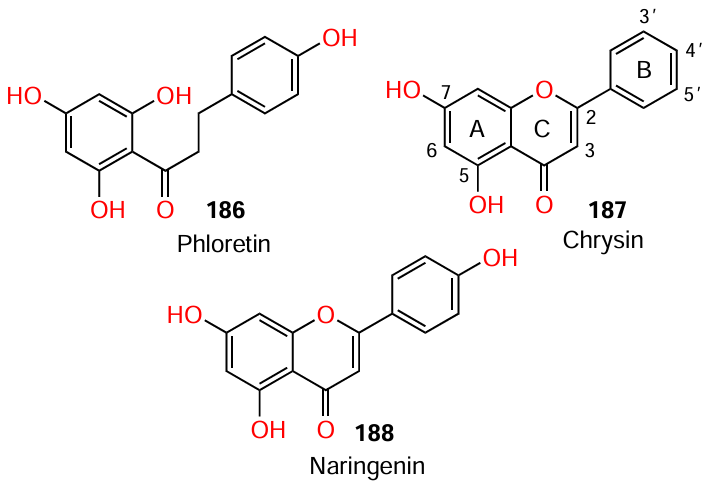
A thorough investigation of the structure – activity relationships for flavonoids revealed the following trends:
(1) the presence of two hydroxyl groups in ring A is indeed important for inhibitory activity; one hydroxyl group must be located at the C(7) atom, while the position of the second hydroxyl group is not critical;
(2) the presence of other hydroxyl groups in ring A is acceptable, but more bulky substituents such as methyl group in ring A or B are undesirable.
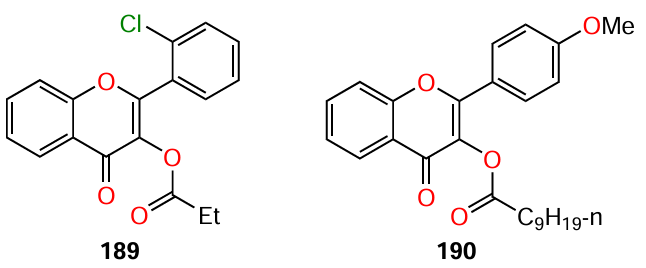
Manner and Fallarero,[122] who searched for effective QS inhibitors, selected two lead compounds 189 and 190, which inhibited the violacein production in C. violaceum ATCC 31532 by > 90 and > 83.5%, respectively, when used at a concentration of 40 mM. It can be seen that the two flavonoids have a structural similarity, in particular, they contain side chains in the acyl substituent of ring C and, hence, they can be regarded as AHL analogues.
12. Fatty acids
As has already been mentioned in the Introduction, DSFs, which are fatty acids produced by bacteria of one species, can suppress competing bacteria of a different species through the bacterial interspecies communication. It is reasonable to assume that other fatty acids of non-bacterial origin may also have anti-quorum sensing activity against Gram-positive and Gram-negative bacteria, owing to their structural similarity to DSFs.
Plants represent a major source of fatty acids, both saturated and (poly)unsaturated ones.[123] For example, industrially valuable safflower oil (Carthamus tinctorius L.) is rich in oleic acid (> 75% of the oil composition).[124] Branched saturated acids, which are most similar to bacterial DSFs, although previously traditionally considered to be of only animal or bacterial origin, have also been found in plants.[125-127] The effect of certain acids with long aliphatic chains (e.g., compounds 191 – 195) on biofilm formation in S. aureus and P. aeruginosa bacteria was studied.[121]
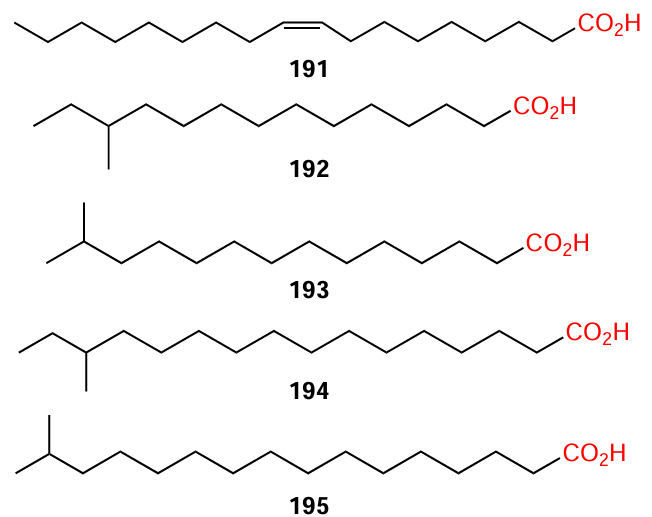
Analysis of the limited data revealed the following aspects of the structure – activity relationship for fatty acids:
(1) for anti-biofilm activity, the acid molecule should preferably contain a double bond; it is desirable (but not necessary) that the double bond is at position 2 (see the formulas of cis-dec-2-enoic and cis-dodec-2-enoic acids in Table 1). For example, oleic acid (191) inhibits biofilm formation in S. aureus and P. aeruginosa exactly by affecting the QS systems;
(2) fatty acids with a chain length of 8 to 14 carbon atoms are the most active;
(3) branched saturated acids such as 12-methyltetradecanoic acid (192) (at a concentration of 10 μg mL–1), 13-methyltetradecanoic acid (193), 14-methylhexadecanoic acid (194), and 15-methylhexadecanoic acid (195), have an inhibitory effect not only on the biofilm formation, but also on the swimming and swarming of P. aeruginosa.
13. Sulfur-containing compounds
Fong et al.[128] synthesized 29 analogues of ajoene (196), an active component of garlic (Allium sativum), known for its antimicrobial properties.[129] In order to establish the structure – activity relationships, the quorum sensing inhibitory activity of the isolated compounds against P. aeruginosa PAO1 was studied. Of the whole series, compounds 197 – 219 exhibited inhibitory effect that was not associated with bactericidal properties, with most of them being more active than ajoene (Table 18). Benzothiazole analogues 198 and 200 proved to be the most active (IC50 = 0.98 ± 0.14 mM and IC50 = 0.56 ± 0.11 mM, respectively).
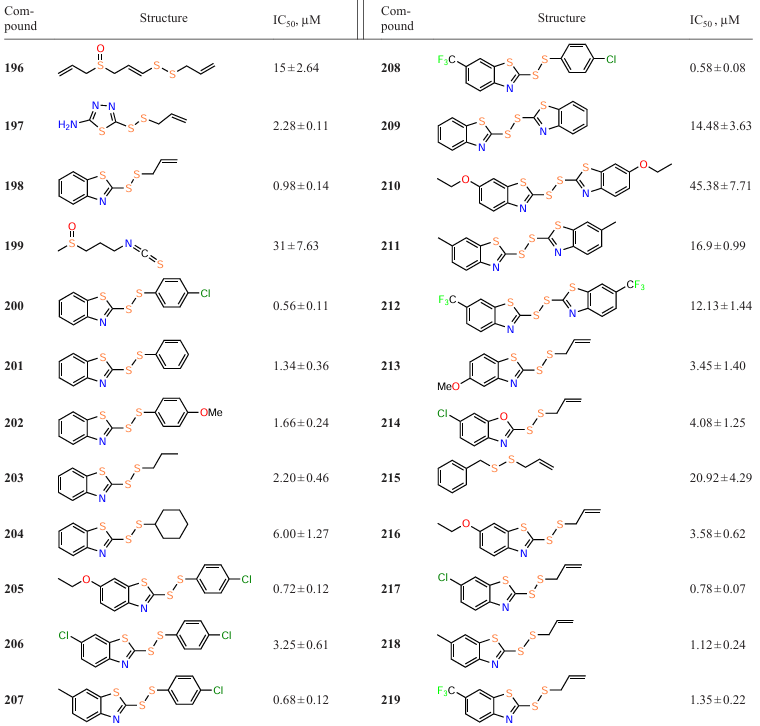
These results provide the following conclusions:
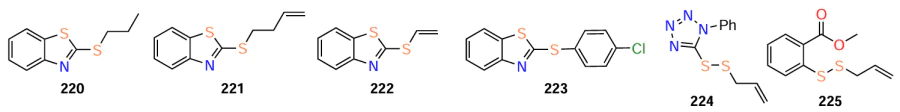
(1) the presence of a disulfide group is an important factor for the quorum sensing inhibitory activity (compounds 220 – 225 proved to be inactive);
(2) conversely, the presence of an allyl group is not essential (saturated analogue 203 proved to be an excellent QS inhibitor);
(3) hydrophobic and bulky substituents decrease the anti-quorum sensing activity (based on the values for compounds with cyclohexyl (204) and benzothiazole (209 – 212) groups);
(4) the introduction of a benzothiazole (both substituted and unsubstituted) or benzoxazole (less preferred) ring leads to a higher activity;
(5) in addition to the disulfide bond and benzothiazole moiety, the introduction of a p-chlorophenyl substituent into the molecule usually leads to an increase in the inhibitory activity, unlike the introduction of a phenyl or p-methoxyphenyl group.
Another readily available and promising sulphur-containing inhibitor of quorum sensing in P. aeruginosa is iberin (226). This compound was isolated from the ethyl acetate extract of horseradish (Armoracia rusticana), a plant that, like garlic, has antimicrobial properties.[130][131] Jacobsen et al.[132] compared the quorum sensing inhibitory action of iberin (226) and its synthetic analogues, isothiocyanates 227 – 230, against two P. aeruginosa strains. However, in this case, the authors were unable to obtain a derivative that would be superior to the natural compound in activity (Table 19).
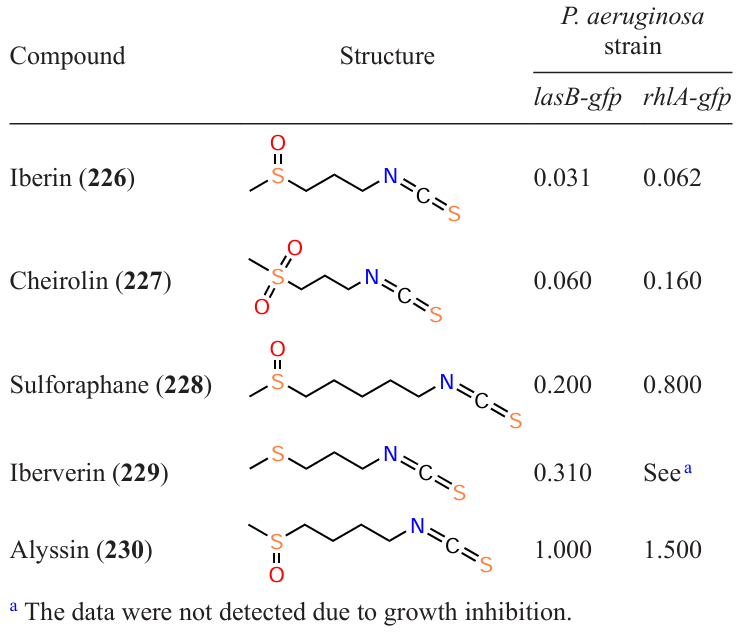
Regarding the structure – activity relationship, the authors made the following conclusions:
(1) in terms of QS inhibition, the presence of a sulfinyl group in the molecule is much more preferable than the presence of a sulfonyl group;
(2) three-carbon chain has advantages over a chain consisting of four or five atoms.
14. Alkaloids
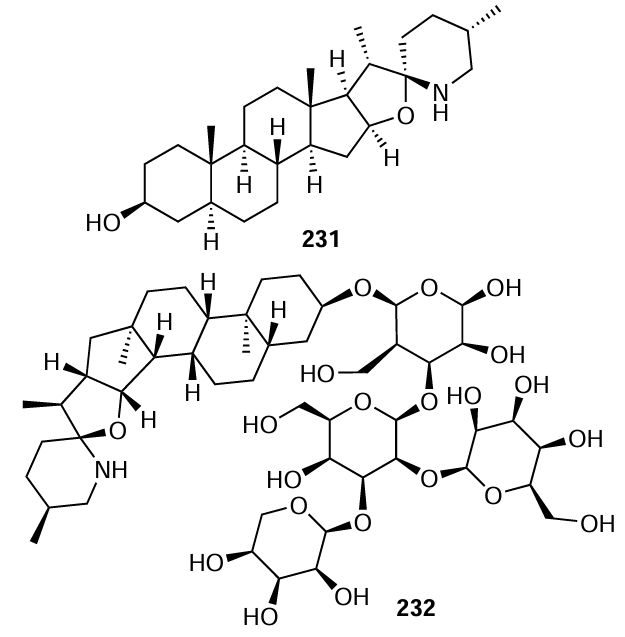
Although the boundary between alkaloids and nitrogen-containing natural compounds (or biogenic amines) is vague, alkaloids are defined, most often, as naturally occurring azaheterocyclic compounds (mainly isolated from plants) that possess basic properties and high physiological activity. Although the antibacterial action of alkaloids is mentioned in a large number of review articles (e.g., a review by Cushnie et al.[133]), there are currently almost no published data on the structure – activity relationship with regard to the QS inhibition for alkaloids. An exception is the paper by Mitchell et al.,[134] who found that tomatidine (231) at a concentration of 12.8 mg L–1 markedly inhibits the hemolytic activity and the expression of agr-regulated genes (hla, hld, geh, nuc, plc, and splC) responsible for the production of virulence factors in various strains of S. aureus. At the same time, its glycosylated derivative, tomatine (232), has virtually no inhibitory effect.[134]
15. Conclusion
This review is the first attempt to summarize scattered data available in the literature concerning the structure – activity relationships for plant-derived quorum sensing inhibitors. The review has certain limitations. For instance, particular molecular targets have not been considered and small samples of compounds were used. Furthermore, due to the lack of information on some secondary plant metabolites, it was necessary to analyse literature data on their synthetic analogues.
Currently, due to the great structural diversity of naturally occurring QS inhibitors, it is difficult to identify the most promising class and to elucidate common trends of the influence of functional groups on the activity. However, the following general features of the structure–anti-quorum sensing activity can be identified:
most QS inhibitors are full or partial structural analogues of either proper AHLs (e.g., γ-lactones, terpenes, zingerone and its derivatives) or functional AHL mimic, that is, rosmarinic acid (e.g., curcuminoids and stilbenes);
for many compounds (stilbenes, flavones, coumarins, and quinones), the number and location of hydroxyl groups is a key structural factor determining the presence and specificity of the quorum sensing inhibition activity.
It should be noted that the prospects of using plant-based QS inhibitors and their derivatives as alternatives to antibiotics and synthetic antimicrobial drugs lies in the fact that most of them have additional types of activity, such as anti-inflammatory, anticancer, antidiabetic, antiviral, antifungal activity, etc.
As regards the future application of the compounds described in this review, the development of new functional materials modified with QS inhibitors appears to be a promising area. These may be catheter coatings that prevent the spread of infection and the growth of bacterial biofilms[135] or films for food packaging that prevent contamination and spoilage of food products.[136][137] The applicability of QS inhibitors as an alternative to food preservatives is also being considered.[138][139] In any case, the practical potential of these compounds has not yet been fully revealed.
This review was written with the financial support of the Russian Science Foundation (Project No. 23-73-10158).
16. List of abbreviations and symbols
The following abbreviations and symbols are used in this review:
AIP — autoinducing peptides,
AHL — N-acetylated homoserine lactones,
AI — autoinducer,
All — allyl,
DPD — 4,5-dihydroxypentane-2,3-dione,
DSF — diffusible signal factor,
EC — effective concentration,
HSL — homoserine lactones,
IC50 — half-maximal inhibitory concentration,
MBC — minimum bactericidal concentration,
MIC — minimum inhibitory concentration,
MRSA — methicillin-resistant Staphylococcus aureus,
PQS — 2-heptyl-3-hydroxy-4-quinolone,
PSM — phenol-soluble modulins,
QS — quorum sensing,
SrtA — sortase A,
T3SS — type III secretion systems,
VRSA — vancomycin-resistant S. aureus.
References